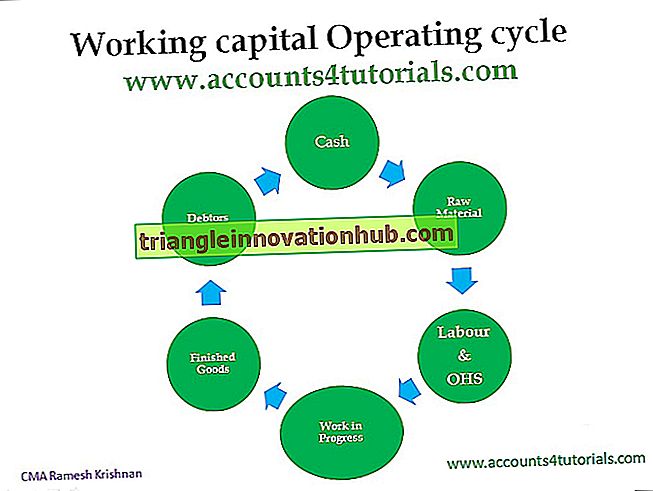
2 خطوات رئيسية تشارك في آلية تخليق البروتين: النسخ والترجمة
الخطوات الرئيسية التي ينطوي عليها آلية تجميع البروتين هي 1. النسخ و 2. الترجمة!
وتخضع عملية التخليق الحيوي للبروتين للسيطرة المباشرة على الحمض النووي في معظم الحالات أو تحت سيطرة الحمض النووي الريبي الوراثي حيث يوجد الدنا.
يتم تخزين المعلومات لهيكل بولي ببتيد في سلسلة polynucleotide. في عام 1958 ، اقترح كريك أن يتم نقل المعلومات الموجودة في الحمض النووي (في شكل تسلسل قاعدة) إلى الحمض النووي الريبي ومن ثم من RNA يتم نقلها إلى البروتين (في شكل تسلسل الأحماض الأمينية) ، وأن هذه المعلومات لا تتدفق في الاتجاه العكسي ، أي من البروتين إلى الحمض النووي الريبي إلى الحمض النووي.
توفر جزيئات الدنا المعلومات لتكرارها. وتعرف هذه العلاقة بين DNA و RNA وجزيئات البروتين بأنها عقيدة مركزية. ذكر تيمين (1970) أن الفيروسات القهقرية تعمل على عكس العقيدة المركزية (التدفق العكسي للمعلومات) أو الحركة النسائية داخل الخلايا المضيفة.
يقوم الحمض النووي الريبي الجيني لهذه الفيروسات أولاً بتجميع الحمض النووي من خلال النسخ العكسي ؛ يتم تحفيز هذه العملية عن طريق إنزيم المنتسخة العكسية ، ثم ينقل الحمض النووي المعلومات إلى الرنا المرسال الذي يشارك في ترجمة المعلومات المشفرة لتشكيل polypeptide.

آلية تخليق البروتين:
(ط) هناك خطوتان رئيسيتان في تركيب البروتين ؛ (1) النسخ ، التي تنطوي على نقل المعلومات الجينية من الحمض النووي إلى الرنا المرسال ، و (2) الترجمة ، التي تنطوي على ترجمة لغة الحمض النووي إلى ذلك من البروتينات.
I. النسخ:
يُعرف نقل المعلومات الوراثية من الحمض النووي إلى الرنا المرسال (mRNA) باسم النسخ (transcription). تقوم بوليميراز RNA واحد بتوليف جميع الرنا (بما في ذلك mRNA و rRNA و tRNA) في البكتيريا. من ناحية أخرى ، تحتوي حقيقيات النواة على ثلاثة بوليمرات مختلفة من الرنا.
تم تعيين واحد من هذه الموجود في النيوكليولز على شكل بوليميراز RNA I أو "A" وهو مسؤول عن توليف rRNA. تم العثور على ثاني بوليمروز الحمض النووي الريبي حقيقيات النواة في النواة ، ويسمى RNA polymerase II أو 'B' وهو مسؤول عن تخليق سلالات mRNA تسمى RNA النووي المتغاير (HnRNA).
تم العثور على ثالث بوليمراز RNA حقيقيات النواة أيضا في النواة ، ويسمى RNA polymerase III أو 'C' وهو المسؤول عن تخليق الحمض النووي الريبي 5S و tRNA. كما تحتوي حقيقيات النواة على بوليمرات أخرى من الرنا في الميتوكوندريا والبلاستيدات.
يتكون بوليميراز الحمض النووي الريبي البكتيرية من أربع سلاسل مختلفة من polypeptide: إنزيم أساسي x (سلسلتين من ∞ وسلسلة مفردة لكل من β و β) وعامل سيغما (a)
1. نسخ من mRNA من الحمض النووي:
في وجود إنزيم بوليميراز الحمض النووي الريبي يعتمد على الحمض النووي يتم نسخ الرسالة الجينية المشفرة في الحمض النووي في مرنا. يعمل فرقتان جزيء الحمض النووي الخاص uncoil وأحد هذين الخيوطين ، كقالب (يطلق على هذا الشريط اسم strins antisense) ، والذي يتم من خلاله نسخ تسلسل دقيق من النوكليوتيدات إلى جزيء mRNA. ونتيجة لذلك ، يكون تسلسل القاعدة لجزيء الرنا المرسال مكملاً لخط الجديلة المضاد للحساسية الذي كان بمثابة قالب. مثل توليف الحمض النووي ، تخليق الحمض النووي الريبي أيضا من 5 ′ إلى 3 ′ الاتجاه (5 ′ - »3 ′).
(أ) النسخ في بدائيات النوى:
في البكتيريا ، يحفز فقط بوليميراز الحمض النووي الريبي وحيد توليف أنواع مختلفة من الرنا. تتكون بوليميراز الحمض النووي الريبي من أربعة سلاسل ببتيدية (αββ'α 2 ) التي تشكل إنزيمًا أساسيًا وعامل سيغما (σ) ، والذي يتم ربطه بشكل فضفاض بالأنزيم الأساسي. يساعد عامل سيغما في التعرف على إشارات البدء على جزيء الحمض النووي ويوجه بوليميراز الحمض النووي الريبي في اختيار موقع البدء. في غياب σ ، يبدأ الإنزيم الأساسي بتخليق الحمض النووي الريبي بطريقة عشوائية.
وبمجرد بدء تركيب الرنا ، ينشأ الإنسحاب والإنزيم الأساسي عن استطالة الرنا المرسال.
آلية النسخ في بدائيات النوى بالتالي تتضمن الخطوات التالية:
1. تجليد RNA polymerase holoenzyme إلى موقع مروج. وقد تم تحديد عدد كبير من هذه المواقع ، معظمها قبل نقطة البدء (أي المنبع) ، ولكن نادرا ما يتم أيضا بعد نقطة البدء (أي المصب).
2. تفكيك الحمض النووي ، مما يؤدي إلى فصل اثنين من فروع والتي يتم نسخ واحد فقط.
3. انفصال عامل سيغما (أ).
4. استطالة نسخة مرنا بمساعدة الإنزيم الأساسي.
5. إن إنهاء تصنيع mRNA يتم عن طريق كودون الإنهاء على DNA. في البكتيريا يتم التعرف على إشارة الإنهاء هذه بواسطة العامل rho (P).

(ب) النسخ من mRNA في حقيقيات النواة:
في حقيقيات النوى هناك نوعان على الأقل من بوليمرات الحمض النووي الريبي. الحمض النووي الريبي بوليميريز- A هو المسؤول عن توليف rRNA. إن RNA polymerase-B تجلب توليف الحمض النووي الريبي النووي (HN-RNA) من الحمض النووي RNA ، وهو عبارة عن تسلسل يتكون من حوالي 200 نيوكليوتيد من حامض adenylic ، أي poly A (poly-adenylic acid) يتم ربطه بـ 3 of end of Hn-RNA. في نفس الوقت يتفكك Hn-RNA عند نهاية 5. ، ويعرف المنتج النهائي باسم poly-A-mRNA.
ينتشر من النواة إلى السيتوبلازم ، حيث يتم استخدامه لتخليق البروتين (يحمل كلا النهايتين تسلسل نوكليوتيدي معين. يمتلك 5 of نهاية جزيء mRNA 7-methyl guanosine ، بينما ينتهي 3 in في تسلسل poly A. إن تسلسل النوكليوتيدات عند طرفي كل جزيئات الرنا المرسال هو نفسه ، لذلك يقال إن جزيئات mRNA لها نهايات ملحوظة.
تشكيل أميناسل - الحمض الريبي النووي النقال:
أظهرت الدراسات التي أجراها <لي Lipمان> وزملاؤه خلال عقد الخمسينيات أن الأحماض الأمينية مرتبطة بجزيئات الحمض الريبي النووي النقال: هذا المرفق له الخطوتين التاليتين:
1. تتكون الخطوة الأولى من تنشيط الأحماض الأمينية ؛ يتفاعل جزيء الحمض الأميني مع جزيء ATP (أدينوسين ثلاثي الفوسفات) لإنتاج جزيء aminoacyl-AMP (amino-acyl adenylate) وجزيء واحد من pryophosphate (PP).

2. في الخطوة الثانية ، يتم نقل الحمض الأميني من جزيء aminoacyl-AMP إلى جزيء معين من الحمض الريبي النووي النقال ويتم تحرير جزيء AMP (أدينوسين أحادي الفوسفات).

يتم تحفيز كل من ردود الفعل من قبل نفس الانزيم ، synthetase aminoacyl-tRNA. يرتبط معقد الحمض الأميني- AMP بإحكام بالأنزيم خلال التفاعل بأكمله. تتفاعل مجموعة الكربوكسيل من الأحماض الأمينية مع مجموعة -OH من بقايا فوسفات AMP لتشكيل aminoacyl adenylates ، في حين أنه يعلق على واحدة من مجموعات -OH من ريبوز من النيوكليوتيدات الأدينين الطرفية لإنتاج aminoacyl-tRNA.
يحتوي كل حمض أميني خاص به على أمينوآسيل tRNA-synthetase خاص به ، وقد تحتوي بعض الأحماض الأمينية على أكثر من إنزيم تنشيط.
II. ترجمة:
تتضمن خطوة الترجمة ترجمة لغة الأحماض النووية (المتوفرة على شكل mRNA) إلى لغة البروتينات.
يمكن تقسيم عملية الترجمة إلى الخطوات المتميزة التالية:
(1) الشروع
(2) استطالة و
(3) إنهاء.
1. بدء سلسلة polypeptide:
إن بدء سلسلة polypeptide يتم إحضارها دائمًا عن طريق methionine الحمض الأميني ، والذي تم ترميزه بواسطة كودون AUG. في E.coli اثنين من الحمض الريبي النووي النقال مختلفة تلقي methionine - (i) tRNA m التقى (tRNA غير قابل للنمو) و (ii) tRNA f التقى (tRNA formylatable). الحمض النووي الريبي النووي الرناوي تودع الميثيونين في شكل أول حمض أميني من سلسلة بولي ببتيد وبالتالي يبدأ تشكيل سلسلة بولي ببتيد. التقى الحمض الريبي النووي النقال م methionine في موقف داخل في سلسلة بولي ببتيد.
يعني كل رسالة تبدأ بـ codon AUG.
(1) بدء سلسلة بولي ببتيد في بدائيات النوى:
في البدائية بدء الشره عن طريق methionine formylated.
في E.coli formylated methionine يتم التقاطه بواسطة الحمض الريبي النووي النقال الذي تم تحديده بواسطة الحمض النووي الريبي النووي (tRNA f met metetal). يتم ترسيب الميثيونين في الموضع الداخلي في سلسلة polypeptide بواسطة آخر tRNA-tRNA met meton (tRNA غير قابل للنمو).

تعلق الميثيونين المصاغة بحمض الحمض النووي الريبي النووي (fR) الذي تم التوصل إليه مع تشكيل f-met-tRNA f . ترتبط الوحدة الفرعية الصغيرة للريبوسوم (30S) بـ5 of طرف mRNA تحمل كودون AUG لتشكيل معقد بدء (30S-mRNA). تم تسهيل ذلك بواسطة عامل بروتين البدء 1F 3 f-met-tRNA f الذي تم ربطه بمركب البدء المكون من 30S-mRNA-f-met-tRNA f metre؛ عامل البدء 1F 2 ضروري لهذه الخطوة. ويجمع هذا مع الوحدة الفرعية الكبيرة (50S) التي تكمل تشكيل ريبوسوم 70S. تستخدم هذه العلاقة الطاقة بسبب انشقاق جزيء GTP واحد.

2. استطالة سلسلة Polypeptide:
بعد تكوين 70S-mRNAf-met-tRNA f met complex ، يحدث استطالة سلسلة polypeptide بواسطة إضافة منتظمة للأحماض الأمينية في الخطوات التالية: -
(1) تجليد الـ AA-tRNA في الموقع- A لوحدة أكبر من الريبوسوم:
من المحتمل أن يكون مركب الحمض النووي الريبي النووي الأميني الأحيائي (AA-tRNA / - Met-tRNA t المتصل بالموقع المستقبلي على الوحدة الفرعية الأكبر للريبوسوم (A- site) ويرتبط الحمض النووي الريبي (tRNA) الحامل لسلسلة الببتيد بموقعه peptidyl أو المانحة (P تتضمن هذه العملية جزيءًا من GTP يوفر الطاقة اللازمة ، ويربط الحمض النووي الريبوزي الأميني (aminoacyl tRNA) الثاني بالموقع A ويربط بالمدونة الثانية لـ GCU على mRNA ، وهو موقع لربط الحمض الريبي النووي النقال.

(2) تشكيل رابطة الببتيد:
يتم تشكيل رابطة الببتيد بين مجموعة COOH من الحمض النووي الريبوزي الببتيديل في موقع- P و- مجموعة NH 2 من aminoacyl tRNA من الموقع A. بعد تشكيل رابطة الببتيد ، يتم تحرير الحمض الريبي النووي النقال من موقع P ويتم نقل سلسلة polypeptide إلى الحمض الريبي النووي النقال الموجود على موقع A.
(3) حركة الحمض الريبي النووي الريبي منقوص الأكسجين peptidyl من موقع A إلى موقع P:
حالما يتم إطلاق الحمض الريبي النووي النقال من P-site ، فإن peptidyl-tRNA من A-site يتحول مرة أخرى إلى موقع P. اكتمال هذه العملية بمساعدة جزيء من GTP وعامل نقل أو انزيم translocase.
خلال هذه العملية ، تتغير الريبوسوم على طول الرنا المرسال في اتجاه 5-3، ، بحيث يتوفر الكودون التالي على mRNA في الموقع A. وهذا يتطلب G-factor و GTP.
عندما يتحرك ريبوسوم واحد على طول مرنا ، تصبح نقطة البدء في mRNA مجانية. يمكن أن تشكل معقد بدء مع وحدة فرعية 30S من ريبوسوم آخر. بهذه الطريقة ، يحصل عدد من الريبوسومات على جزيء مرنا وحيد. يعرف هذا المجمع باسم مجمع polyribosome.
خلال عملية تخليق البروتين ، يمكن رؤية عدد من الريبوسومات مرتبطة بجزيء مرنا وحيد في وقت واحد ، مع كل سلسلة من polypeptide تحت التكوين ، يكون حجم سلاسل polypeptide على ريبوسوم مختلفة مختلفة.
يوجد عامل بدء آخر IF1؛ وهو أصغر عوامل البدء الثلاثة (IF1- 9500 daltons و IF2 و 73.000 daltons و IF3 = 23000 daltons) ولا يتم فهم دوره بوضوح. قد يكون مهتمًا بالمساعدة في إصدار IF2 من مجمع الاستهلال.
(4) بدء سلسلة polypeptide في الكائنات حقيقية النواة:
في حقيقيات النواة ، يتم استهلال بداية سلسلة من polypeptide بواسطة met-tRNA خاص ، لكن الميثيونين غير مهيأ (لأن الحمض النووي الريبي النووي الذائب لا يوجد في النباتات ، و transylcise الإنزيمي غائب في الحيوانات). في حقيقيات النوى ، وحدة أصغر (40 S) من الزميلة الريبوسوم مع الحمض الريبي النووي المبتدئ المعروفة باسم tRNAy f met .
40S + Met-tRNA التقيت 40S- اجتمع Met-tRNA
40S - Met-tRNA + mRNA -> 40S-mRNA-met- tRNA met
40S - mRNA-met-tRNA i met + 60S -> 80S -mRNA-met- tRNA i met
يوجد في حقيقيات النواة ما لا يقل عن عشرة عوامل بدء مختلفة. هذه هي elF1 و eIF2 و eIF3 و eIF4A و eIF4B و eIF4C و eIF4D و eIF4F و eIF5 و eIF6. يشبه eIF3 و eIF2 IF2 و IF3 من بدائيات النواة.
3. إنهاء Polypeptide:
ويحدث ذلك من خلال وجود أي من أكواد الإنهاء الثلاثة ، وهي UAA و UAG و UGA. يتم التعرف على هذه الكودونات إنهاء بواسطة واحد من اثنين من عوامل الإصدار RF1 و RF2 في E.coli. من بين عوامل الإصدار هذه ، يتعرف RF1 على UAA و UAG ، بينما يتعرف RF2 على UGA و UAA. إنها تساعد الريبوسوم على التعرف على هذه الثلاثية.
يبدو أن عوامل الإطلاق تعمل على A 'لأن الحمض الريبي النووي النقال القامع القادر على التعرف على كودونات الإنهاء يمكن أن يتنافس مع عوامل الإطلاق عن طريق الدخول في الموقع A. يبدو أن عوامل الإصدار الثالث RF3 تحفز عمل RF1 و RF2.
بالنسبة لرد فعل الإطلاق ، يجب أن يكون polypeptidyl tRNA موجودًا في موقع "P" وتساعد عوامل الإطلاق في تقسيم مجموعة الكربوكسيل بين عديد الببتيد و tRNA الأخير الذي يحمل هذه السلسلة. وبالتالي يتم إطلاق البولي ببتيد وينتسخ الريبوسوم إلى وحدتين فرعيتين بمساعدة IF-3.
في eukaryote يُعرف عامل إطلاق واحد فقط ، أي eRF1.
4. تعديل سلسلة Polypeptide المفرج عنها:
يتم إزالة مجموعة الفورمالين من أول حمض أميني ، ميثيونين ، من سلسلة بولي ببتيد تم إطلاقها بواسطة إنزيم ديفورميلاز. بعض الانزيمات الأخرى مثل expeptidases تزيل بعض الأحماض الأمينية إما من طرف الطرف N أو طرف الطرف C أو من طرفي سلسلة polypeptide. وأخيرًا ، يتم طي سلسلة البولي ببتيد هذه بمفردها أو جنبًا إلى جنب مع السلاسل الأخرى لتحمل البنية الثلاثية أو الرباعية وتعديلها إلى إنزيم وظيفي.