قسم الخلية: Amitosis ، الانقسام الخلوي ، السيتوبلايزيس
قسم الخلية: Amitosis، Mitosis، Cytokinesis!
هناك نوعان من الكائنات الحية - الخلوية و متعددة الخلايا. يعتمد نمو وتطور الفرد حصريًا على نمو الخلايا وتكاثرها. لقد كان فيرتشو هو أول من أعلن عن تقسيم الخلية.
في الخلية الحيوانية تمت دراسة الانقسام الخلوي على شكل تقسيم التقسيم أو الانقسام بواسطة Prevost و Dumas في عام 1824. لم يتم التحقيق بدقة في آلية الانقسام الخلوي إلا بعد مرور فترة طويلة ولكن أظهرت Remak و Kolliker أن العملية تتضمن تقسيم كل من النواة والسيتوبلازم.
تم إدخال مصطلح karyokinesis بواسطة Schleicher (1878) لتعيين تغييرات النواة أثناء الانقسام ، وتم إدخال مصطلح cytokinesis بواسطة Whiterman (1887) لتحديد التغيرات المصاحبة التي تحدث في السيتوبلازم.
تقسيم الخلايا هو بالضرورة تجنب الشيخوخة وثانيا لفصل الفرد إلى وحدات شبه مستقلة مما يؤدي إلى الكفاءة. وهكذا ، نرى أن الانقسام الخلوي هو ظاهرة واسعة الانتشار لا تقتصر على الحفاظ على الحياة فحسب ، بل وأيضاً لتطوير الكائن نفسه.
يمكن وصف انقسام الخلية على النحو التالي:
(ط) التقسيم المباشر :
حيث تخضع النواة وجسم الخلية لعملية تقسيم كتلة بسيطة إلى قسمين. ويسمى أيضا البعد.
(2) التقسيم غير المباشر :
هنا تخضع النواة لتغييرات معقدة قبل أن تنقسم إلى نواتين ابنة.
انشطار:
البياض أو الانقسام الخلوي المباشر هو وسيلة للتكاثر اللاجنسي في الكائنات الحية غير الخلوية مثل البكتيريا والأوليات ، وأيضا طريقة لتكاثر أو نمو الأغشية الجنينية لبعض الفقاريات. في نوع انزيمات الخميرة ينقسم تقسيم النواة إلى انقباض السيتوبلازم.
أثناء نقع البُقَى ، تستمد النواة أولاً ثم تفترض المظهر على شكل دمبل. يزداد حجم الاكتئاب أو الانقباض ويقسم النواة في النهاية إلى نواتين ؛ ويتبع تقسيم النواة انقباض السيتوبلازم الذي يقسم الخلية إلى نصفين متساويين أو متشابهين تقريبًا.
لذلك ، دون حدوث أي حدث نووي يتم تشكيل خليتين ابنة.

الانقسام المتساوي:
في الانقسام الفتيلي ، تنقسم خلية واحدة إلى قسمين ، كلاهما متطابقان جينيًا مع بعضهما البعض وإلى الخلية الأبوية. وبعبارة أخرى ، كلاهما ، الكروموسومات والجينات ، هي نفسها في كل الخلايا. هذا النوع من الانقسام الخلوي ضروري إذا كان الكائن الحي و / أو الخلية هو الاستمرار والبقاء.
هناك العديد من الحقائق للحاجة إلى الانقسام الخلوي ، وهي تختلف حسب الوظيفة البيولوجية المحددة. على سبيل المثال ، في أنسجة الكبد عندما تموت أو تتلف بعض الخلايا ، ينقسم آخرون ويقدمون خلية جديدة لتجديد الخلايا المفقودة.
تنمو خلايا أخرى في الكائن الحي بالفعل (زيادة في الحجم) ومن المحتمل أنه عندما تصل إلى نقطة حيث يوجد الكثير من السيتوبلازم بعيدًا عن كمية معينة من المواد النووية ، فإنها تنقسم وتبدأ العملية بأكملها مرة أخرى. تتضمن ظاهرة النمو أيضًا زيادة في عدد الخلايا. الزيادة في حجم النسيج أو العضو غالباً ما تكون بسبب الزيادة العددية في الخلايا وليس زيادة حجم الخلية.
عند إخضاعها للإشارات البيئية والبيوكيميائية المناسبة ، يمكن تحفيز هذه الخلايا للتمييز حتى نوع خلية معين. المجموع الإجمالي هو أنه نتيجة لجميع التقسيمات ، يتم توفير درجة معينة من اللدونة والخلود إلى الكائن الحي.
عندما تُفقد اللدونة ، يخضع الكائن الحي لعملية الشيخوخة ، ولكن عندما تكون عملية التقسيم خارج السيطرة ، فإن الكائن الحي "ينمو" حرفياً حتى الموت! إن عملية الانقسام الفتيلي هي عملية مستمرة ، ولكن على أساس تغييرات معينة يمكن تقسيمها إلى عدة مراحل أو مراحل.
الميزات المميزة للمراحل أو المراحل السحرية :
في الانقسام الخلوي ، تحدث التغييرات المرئية الأولى في النواة ، عندما تتكاثف الكرومونات في الكروموسومات (Gr ، الأجسام الملونة). هذه المرحلة تسمى الطور الأولي (Gr ، الشكل المبكر). ثم يختفي الحد النووي ويصطف الكروموسومات في أو بالقرب من مستوى واحد في الخلية. هذا هو Metaphase (غرام ؛ الرقم الأوسط).
بعد ذلك ، ينفصل كل صبغي إلى جزأين ، وينتقل هذان الجزءان عن بعضهما البعض إلى نهايات الخلايا. هذا هو Anaphase (Gr ؛ صعودا وهبوطا الشكل.). بعد ذلك ، تعيد الكروموسومات عند كل نهاية الخلية إعادة تكوين نواة. هذا هو Telophase (غرام ، الشكل النهائي). ثم ينقسم السيتوبلازم إلى خليتين ، مما يجعل الطور البيني مرة أخرى.
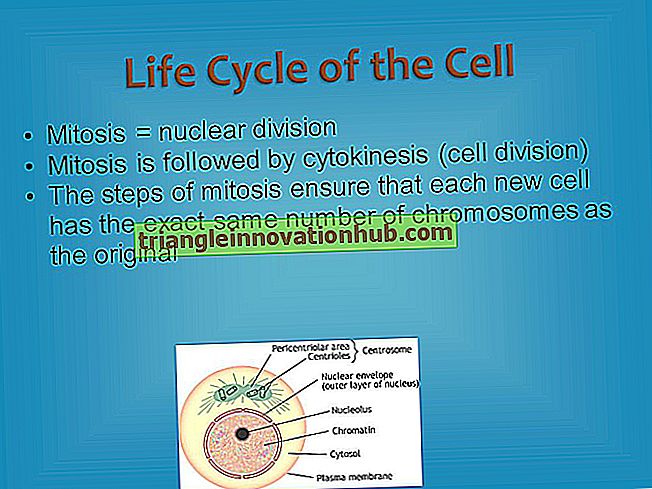
البيني:
وقد سميت فترة النشاط الأيضي خلال أي عملية الانقسام الخلوي في عملية "المرحلة البينية". غالباً ما يشار إلى هذا باسم "مرحلة الراحة" ولكن هذا المصطلح غير مناسب لأن الخلية هي الأكثر نشاطاً في عملية التمثيل الغذائي في هذه المرحلة ولهذا السبب أشار Berril و Huskins (1936) إلى أنها مراحل الطاقة.
المرحلة البينية هي الفترة بين الطور البالي لقسم واحد و الطور الأول لقسم الخلية الجديد. خلال هذه المرحلة ، تقوم الخلية بكل شيء باستثناء القسمة. فمن خلال الطور البيني أن الجينات مكررة ذاتياً وتتابع وظيفتها في الإشراف على التوليف.

إن حبيبات الكروماتين في النواة لا يمكن تمييزها بسهولة في الخلية الحية ، ولكن يمكن إخراجها عن طريق المعالجة باستخدام مادة كيميائية تقتلها وتثبتها وتلصقها. وهي تظهر للوهلة الأولى أنها مبعثرة في جميع أنحاء النواة ولكن الدراسة المتأنية قد أنتجت دليلاً على أنها مرتبة في نمط محدد كسلاسل طويلة ملفوفة وتظهر على هيئة خيط مثل العمل الصافي في التحضير العادي الملون.
"الانفعال النووي" أو "Karyoplasm" يملأ الفجوات بين الكروموسومات. واحدة من أكثر الهيئات المستديرة ، وعادة ما تكون موجودة النواة. بين النواة والسيتوبلازم المحيط بها ، هو الغشاء النووي. في السيتوبلازم المتاخم للنواة ، يوجد جسم ، الجسم المركزي ، يتألف من حبيبين ، أو بعد كل حبيبة قد تكررت ، من زوجين من الحبيبات ، السيقان الوسطى.
دورة الخلية النموذجية ، بما في ذلك الطور البيني تستمر من 20-24 ساعة. المرحلة البينية هي أطول فترة في دورة الخلية وقد تستمر لعدة أيام في الخلايا.
يمكن تقسيم interphase كذلك إلى أربع مراحل فرعية:
1. ز 1 - المرحلة.
2. S- المرحلة.
3. ز ، المرحلة.
4. M-Phase.
يتضمن G1 مرحلة تركيب وتنظيم الركيزة والانزيم ، اللازمة لتخليق الحمض النووي. لذلك ، فإن G1 ، يتميز بتخليق RNA والبروتين. G 1 - المرحلة تليها المرحلة S حيث يحدث تخليق الحمض النووي. خلال G 2 - المرحلة ، يتم تنفيذ جميع الأنشطة الأيضية. المرحلة M هي فترة الانقسام الكروموسومي.
الأطوال النسبية لهذه الأطوار تختلف في الكائنات الحية المختلفة. وتكمل الخلية البشرية في الثقافة عند 37 درجة مئوية ، الدورة الانقسامية في حوالي 20 ساعة وتستمر المرحلة M لساعة واحدة فقط. تلعب درجة الحرارة وبيئة الخلية دورًا مهمًا في تحديد معدل انقسام الخلايا.
يمكن حتى في بعض الأحيان أن يتم تقسيم الخلايا غير المورفستولوجية من خلال تغيير الظروف البيئية. تلك الخلايا التي لن تنقسم بعد الآن ، لديها دورة الانقسامية في مرحلة G : وتبدأ بالتمييز.
تظهر الخلايا التغييرات التالية في الطور البيني:
1. الخلية ، ككل ، تبلغ أقصى نمو وتمتلك البروتينات المركبة للطاقة ، لمختلف التقسيمات والعمليات.
2. الغشاء النووي سليم ويتم العثور على الكروموسومات في شكل خيوط ملفوفة بشكل أو بآخر بشكل أو بآخر ، ومشبعة إلى حد ما بالغشاء. في هذه الحالة من الكروموسومات ، فإن معظم علماء الخلايا يعتبرونها مزدوجة ، بينما يرى بعض العمال أنها متعددة الأجزاء.
3. اثنين centrioles ، التي توجد في الزوايا الصحيحة لبعضها البعض تتكاثر إلى اثنين لكل منهما. وقد وصف Mazia (1961) أنه إذا تم التحقق من النسخ المتماثل ، فلن يحدث الانقسام.
4. بالنسبة لتكثيف المغزل المستقبلي لل protروتوبلازم في منطقة متماسكة من الاتساق الشبيه بالهلام يحدث أيضاً. يبدأ المغزل أيضا في النمو ويدفع الوسط بعيدًا.

6. مركز كرومو أيضا بارز خلال المرحلة البينية.
الطور الأول:
الطور الأولي هو الأطول في M-phase وقد يستغرق حوالي ساعة إلى عدة ساعات. في الخلايا العصبية من جندب يستغرق حوالي 102 دقيقة.
التغييرات الهامة المختلفة في هذه المرحلة هي كما يلي:
1. تميل الخلية إلى أن تصبح كروية وتزيد من انكسارها وثورتها عن طريق زيادة شدتها السطحية عن طريق إزالة الماء.
2. إن النواة تشرب الماء من السيتوبلازم وتندمج الكروموسومات بشكل مختلف. يظهر كل كروموسوم بنيته الغريبة مع عزل ملحوظ للكروماتيدات ، كل منها يمر بدورة منتظمة من اللف.
في بداية الانقسام ، تبدأ الكروموسومات بالتقلص ، ثم تصبح سميكة ، وتخضع لعملية اللف. جزئيًا يبدو هذا التغيير مرتبطًا بالتطوير ، في زوايا قائمة على الدواليب القديمة للملفات. أشار سبارو (1941) إلى أن الانقباض الكلي للختام النهائي هو تقريباً خمس الطول الأولي. من الناحية الفيزيولوجية ، يتم تشكيل اللفات بسبب التكثيف المستمر. الملفات هي من نوعين: ملفات صغيرة أصغر ولفائف جسدية أكبر.
يتم أيضًا تصنيف نمط التواء أثناء تشكيل الملفات والدوائر في صنفين مختلفين:
(ط) Plectonemic:
ملتوية بطريقة لا يسهل عزلها ، و
(2) الوباء:
يمكن فصل الكروماتيدات الملفوفة بشكل جانبي.
3. ترتبط الكروماتيدات مع centromere. يتم توزيع الكروموسومات بشكل منفصل في التجويف النووي. محتويات RNA و phospholipids تزيد تدريجيا.
4. تتعطل الحدود النووية ، ويبدأ جهاز المغزل بالتشكل ، و تختفي النواة و centromere بشكل عام.
يحدث تشكيل المغزل بطريقتين ، وهما:
(1) يقسم المركز الواحد إلى اثنين من centrioles ابنة وعلى الانفصال ، تظهر أشعة نجمي والشعيرات الحساسة ، ودعا المغزل. يهاجر centrioles جنبا إلى جنب مع زهور النجمة حتى تصبح موجودة في المواقف التناقضية. يشار إلى هذا النوع من المغزل على المغزل المركزي.
(2) يتم استقطاب اثنين من centrioles بالفعل قبل بداية التقسيم وتشكيل المغزل يحدث في الطورية. يعرف هذا النوع من المغزل بالمغزل الميتافيزي
ويطلق على الانقسام الفتيلي الذي يتم فيه تشكيل الشكل المغزلي والمغزل من قبل المراكز باسم "الانقسام الفسيولوجي amphiastral" وحيث لا توجد مراكز ، فإن الانقسام الفتيلي يسمى "نجمي". يحدث الانقسام الفلكي في النباتات.
الموالية للالطورية:
هذه المرحلة تتبع الاختفاء الكامل للغشاء النووي. تميل الكروموسومات إلى التجميع في موضع مركزي في الخلية ، بالقرب من خط الاستواء. يتم وصف مصطلح prometaphase بواسطة Coin (1964). وقد عرّفها وايت (1963) بأنها الفترة التي يتم خلالها تشكيل المغزل والتي تعطي خلالها الصبغيات الانطباع بأن بعضها يكافح ويتصارع بعضها البعض في محاولة للوصول إلى خط الاستواء في تطوير المغزل.
على الأقل ، في النباتات ، تتطابق هذه المرحلة مع المظهر الأول لمغزل منظم. اعتبر ويلسون وهايبو (1955) هذا الموقع من الكروموسوم للعب دور مهم ، سواء في التطوير والتنظيم الوظيفي. ألياف المغزل هي أنبوبي ، مرنة ، ليفية وبروتينية في الطبيعة.

الطورية :
يتم تعريف الكروموسومات الطورية بشكل حاد وأجسام منفصلة وتكون ملفوفة بإحكام. في هذه المرحلة ، يمكن عد رقم الصبغي بسهولة ، ومن الممكن التعرف على الكروموسومات المختلفة من أحجامها وشكلها وبنيتها الإجمالية. في الشكل المبين في الطور المبكرة يظهر الشكل المغزلي في المنطقة النووية الواضحة.
يتكون هذا من إشعاعات ليفية تمتد من الجزء المركزي العريض المسمى خط الاستواء وتلتقي عند نقطتين على الأطراف المقابلة للمنطقة النووية ، تسمى الأقطاب. وتبدأ الكروموسومات التي انتشرت حتى الآن بطريقة عشوائية في الجزء النووي المركزي للخلية في إظهار حركات غريبة وترتيب نفسها في منتصف الطريق بين قطبي المغزل ، لتشكيل صفيحة استوائية.
ولكي يكون أكثر دقة ، فإنه يربط بين كرومروم الكروموسومات التي تصطف على الصفيحة الاستوائية. عادة ، تحدث الصبغيات الأصغر بالقرب من مركز الصفيحة الاستوائية والأكبر منها بالقرب من الأطراف الخارجية. ومع ذلك ، ليس من الضروري أن يكون كل من هذين المتناظرين قريبين من بعضهما البعض في اللوحة الاستوائية.
موقف كل كروموسوم مستقل عن الآخر على الصفيحة الاستوائية. الكروموسوم الطوري هو بنية مزدوجة بشكل واضح وينظر بوضوح إلى أن ينقسم طوليًا إلى صبغيين متماثلين تمامًا.
يلعب السنترومير دورًا أساسيًا في تطوير المغزل وفصل كروموسومات الابنة. يتم ترتيب الكروموسومات على خط الاستواء بحيث أن كروماتيد واحد من كل كروموسوم يواجه قطبًا واحدًا والآخر يواجه القطب المعاكس.
ترتبط ألياف مغزلتين بمركز السنتروم لكل كروموسوم ، أحدهما على جانبيها. هذه تربط الكروموسوم إلى القطبين المضادين للمغزل وتسمى بالكروموسوم أو الألياف اللمسية. تمتد الألياف الأخرى للمغزل من القطب إلى القطب ولا ترتبط بالكروموسومات. وتسمى هذه ألياف المغزل المستمر. وتتكون ألياف المغزل أساسا من البروتين ، وبعض حمض الريبونوكليك (RNA) والدهون.
شكل وترتيب الكروموسومات :
ليس ترتيب الكروموسومات عند خط استواء المغزل من نوع واحد في جميع الكائنات الحية ، لأن الترتيب يعتمد على شكل وحجم وعدد الكروموسومات التي تختلف في الكائنات الحية المختلفة ، لأنه في بعض الكائنات الحية تكون (الكروموسومات) خيطية مثل (Urodela). و في الحشرات (Orthoptera و Diptera) ، في Odonata ، Coleoptera و Hamiptera تكون أقصر أو قضيب في شكل Orthopoda يتم تقريبها.
على أساس دراستهم المورفولوجية ، يمكن تقسيمها إلى ثلاث فئات:
(ط) قضبان أو خيوط مستقيمة:
تنشأ هذه الصبغيات مباشرة عن طريق تقصير الخيط spireme.
(ثانيا) الحلقات ، أشكال V أو هوك:
تتشكل هذه الصبغيات بواسطة ثني عند النقطة الوسطى أو بالقرب من أحد الطرفين.
(ثالثا) أشكال حويصلة أو كروية:
وتنشأ هذه الأشكال عن طريق تقصير الخيوط المدقع. جميع الأشكال الثلاثة المذكورة أعلاه هي مزدوجة في الطورية بسبب الانقسام الطولي. يمكن تتبع التقسيم الطولي بين الصبغيات ذات الشكل القضيبي والخيطي بوضوح ، ولكن في أشكال كروية ، غالباً ما يظهر على شكل تضيق عرضي على ما يبدو بسبب التقصير الشديد في الصبغيات.
ارتباط المغزل بالكروموسومات:
نمط ارتباط الكروموسومات بألياف المغزل بناءً على بنية الكروموسومات. ولكن على الرغم من ذلك ، فإن الترتيب وطريقة الارتباط ثابت لكل صبغي معين ، وأنه موروث من جيل إلى جيل. ترتبط الكروموسومات بألياف المغزل بواسطة centromeres. الكروموسومات التي تفتقر إلى ذيل السنترومير لتعلقها بألياف المغزل.
قد تكون طريقة تعلقهم من نوعين:
(ط) محطة أو مركزية.
(2) غير الطرفية أو غير مركزية.
في الوصلة الطرفية ، قد يكون للكروموسومات ارتباطها بألياف المغزل في النهاية الحرة. قد يكون التعلق غير الطرفي في النقطة الوسطى (متوسط) أو عند نقطة وسطية فرعية أو طرف فرعي نقطة intermdiate.

طور الصعود:
في مرحلة ما قبل الطور ، تتحرك أزواج القسيم المركزي على امتداد المغزل وتحمل صبغيًا واحدًا من كل زوج إلى أقطاب متقابلة. المغزل ينمو في النهاية لفترة أطول. حركة الكروماتيد معقدة.
يحدث الفصل الأول للكروماتيد من مركز Centromere ثم يحرك تدفق التيار على طول المغزل حركته القطبية. هذه المرحلة تدوم لفترة قصيرة جدا ، تتراوح بين 6 إلى 12 دقيقة. في المرحلة الأخيرة من الطور ، تزداد المنطقة تدريجياً بين مجموعتي الكروموسومات أو المنطقة الاستوائية. يبدو أن الألياف تمتد وتسمى ألياف interzonal.
وقد يشار إلى expanison من الجزء الأوسط باسم stemmkorper أو دفع الجسم بواسطة الدب. يبدو الجسم المدفوع مثل الجل الذي يدفع مجموعات الكروموسومات نحو الأقطاب المعنية. خلال الحركات القطبية ، تفترض الكروموسومات أشكال "J" أو "V" غريبة ، تبعاً لمركز centromere. في هذه المرحلة ، يُطلق على "J" تسمية heterobranchials و "V'chromosomes as aorobranchials".
حركة الكروموسومات :
يتم التحكم في حركة الصبغي بواسطة ألياف المغزل. في الواقع قد تكون عمليتان تعملان. توسيع مستمر واستطالة المغزل وتقصير الألياف الصبغية. وبما أن الألياف تقصر دون أن تصبح أكثر سمكًا ، فقد تتضمن العملية إزالة الماء أو الجزيئات الأخرى من الألياف. غالبًا ما يُنظر إلى عصبة من "الألياف المتداخلة" لفترة ما بعد انتهاء عملية الانفصال ، مما يربط بين الكروموسومات التي تفككت ، وغالبًا ما تتضمن بقايا المغزل.

القوى المشاركة في حركة الكروموسومات:
ألياف المغزل هي المسؤولة عن حركة الكروموسومات أثناء الطور ، أي من خط الاستواء إلى القطب. تم اقتراح عدة نماذج من قبل عمال مختلفين لشرح القوى المشاركة في الحركة.
بعض منهم يرد أدناه:
(ط) نموذج الانكماش البسيط:
اقترح فان بندن (1883) أن الكروموسومات في الخلية المقسمة يتم سحبها نحو القطبين بواسطة تقلص ألياف المغزل. وصف Swann (1962) أن centromere يفرز بعض المواد التي تسبب في تعاقد أيون ألياف المغزل.
إن الاعتراض الرئيسي على النظرية يأتي من الملاحظات المباشرة على الكروموسومات التي يتم حملها حتى خارج كونترول الذي ترتبط به ألياف المغزل. وعلاوة على ذلك أثناء انقسام الخلايا ، تستطيل الخلية بأكملها والتي هي عكس نموذج الانكماش.
(2) نموذج التوسع:
واقترح Watase (1981) أن ألياف المغزل تمارس الضغط على النواة والصبغ تصبح مسطحة على لوحة الطورية. تصبح الألياف الصبغية مرتبطة بالكروموسومات ، والتي يتم دفعها الآن نحو القطبين المعاكسين. لهذا السبب ، يُعرف هذا أيضًا باسم دفع النموذج.
(3) نموذج الانكماش والتوسع:
اقترح <بيلار> أن الفصل الأولي من كروموسومات ابنة هو عملية مستقلة ، لكن الانفصال الإضافي ناتج عن تقلص أو توسيع أجزاء مختلفة من المغزل. الألياف الصبغية التي تمتد من قرون الكروموسومات إلى أقطاب عقد المغزل وتسحب الكروموسومات المرفقة في القطبين. تتوسع الألياف المشتركة بين المنطقة الموجودة بين الكروموسومات المنفصلة وتدفع كروموسومات ابنة نحو أقطاب متقابلة.
(4) نموذج ديناميكي للتوازن:
يقدم هذا التفسير الأكثر إقناعاً حول الآلية المحتملة لحركة الكروموسومات. وقد وصف Invoke و Sato (1967) حدوث توازن بين برك كبيرة من المونومرات التي تشكل بروتينات الأنيبيبات الميكروية microtubules. خلال البلمرة ، تحدث بعض التفاعلات الكارهة للماء بين المجموعات غير القطبية من مونومرات البروتين.
خلال حركة الكروموسومات يحدث كل من انكماش واستطالة ألياف المغزل عن طريق طرح أو إضافة زوائد أحادية جديدة للألياف. خلال الأطوار الجيوب الأنفية الصبغية ينقبض بحذف مونومير من الطرف القطبي للألياف والألياف المتواصلة تزيد حجم م عن طريق تجميع المواد الجديدة في النهايات القطبية. وبالتالي فإن المغزل يطيل المسافة بين القطبين.
(5) نموذج مزلق:
ويرى Bajer Ostergen وآخرون أن حركة الصبغيات هي عملية نشطة تنساب فيها ألياف الكروموسومات على شكل مراكب شراعية بين الألياف المستمرة. اقترح أمبروز أن القوى المتحركة لهذه الحركات يمكن أن تكون electroshorosis و electrophoresis. من المفترض كسر ATP لتوفير الحجم المطلوب لحركة المغزل.

وصف Mclnuosh و Helper و Van Wie وآخرين وجود جسور ميكانيكية بين الأنيبيبات الدقيقة المشابهة لاتصالات السقاطة في ألياف العضلات. تنساب ألياف الكروموسومات بين الألياف المستمرة وتفرز الكروموسومات. يتم توفير الطاقة عن طريق انهيار ATP.
(7) النموذج الكهربائي:
اقترح Lillie and С oiler (1936) أن التغيير في إمكانات الغشاء يرجع إلى التغيرات المحلية في النفاذية تحدث بالقرب من القطبين وحول الغشاء النووي وتنتج الحقل الكهربائي. تكون الصبغيات مشحونة سلبياً في الطور الأولي ، كما أن الكروموسومات المشحونة تهاجر بسهولة في المجال الكهربائي.
(8) نموذج الدفع :
وفقا لهذا المفهوم يتم دفع الكروموسومات عن طريق المكون الغرواني لخلية السيتوبلازم. يتشرب الماء الغرواني الماء ، ويتضخم ويدفع الكروماتين من الكروموسومات. تعمل ألياف المغزل كمسارات توجه حركة الصبغيات إلى القطبين ، مما يمنعها من التشتت في جميع أنحاء الخلية.
الطور النهائي:
المجموعتين على الكروموسومات مجمعة في القطبين وعند وصولها تفقد تدريجيا لونيتها. يحدث Decondensation مكان. يحدث الإصلاح على الغشاء النووي من خلال عملية مجهولة معينة.
ربما يتم تصنيع المادة الجديدة بواسطة الحمض النووي الريبي خلال الطور البعدي أو ربما يكون من الممكن أن يكون النظام الشبكي الداخلي (endoplasmic) قد أدى إلى ظهور غشاء جديد حول الكروموسومات. تصبح كل الكروموسومات غير مثبتة.
يتم امتصاص ألياف المغزل في منظم السيتوبلازم النووي أو منطقة SAT مرة أخرى تشكل النواة. وأخيراً ، يؤدي هذا التغيير إلى نواتين ابنتين تعادلان النواة الأم في جميع النواحي.
يظهر:
السيتوبلايزيس في النباتات:
يتبع التقسيم النووي أو الانقسام الفتيلي ، كما يطلق عليه ، تقسيم السيتوبلازم. في حين يتم تنظيم نوى ابنة في القطبين ، يختفي المغزل الإنقسامي إلا في خط الاستواء ، حيث تصبح ألياف المغزل المستمر أكثر كثافة.
هذه المنطقة تسمى الآن phragmoplast. وفقا لبورتر و Machade (1960) بدأ تشكيل لوحة الخلية عن طريق هروب العناصر الأنبوبية للشبكة الإندوبلازمية نحو المنطقة الداخلية للمغزل حيث تنتشر لتشكل شبكًا قريبًا على طول الخط الأوسط.
تظهر القطيرات في phragmoplast وتحتوي على مواد peptic التي تندمج لتكوّن في منتصف الخلية المنقسمة قسما يعرف بلوحة الخلية. تضاف قطرات التحريك إلى صفيحة الخلية لتشكيل الصفيحة الوسطى التي تبدأ الآن في التمدد للخارج حتى تصل إلى الجدران الخارجية لجسم الخلية الأساسي للخلية الأصلية المودعة الآن على جانبي الصفيحة الوسطى. يتم تشكيل هذا واثنين من الخلايا. يطلق على تقسيم السيتوبلازم السيتوبلازم.

السيتوبلازمية في الخلايا الحيوانية:
يبدأ التجلط السيتوبيني عند ظهور ثلم ضحل 111 السيتوبلازم عند خط استواء المغزل. ببطء وبطيء تعمق الأخاديد ويضيق السيتوبلازم والخلية إلى بنتين. وقد اقترح عدد من النظريات لتشكيل ثلم.
هذه هي كما يلي:
1. نظرية الحلقة المتعاقبة :
وفقا لسوان وميتشيسون (1958) فإن السيتوبلازم حول خط استواء المغزل يحتوي على بعض البروتينات المقلوبة. تشكل هذه البروتينات نوعًا من الخواتم عند خط الاستواء. مع استمرار انقسام الخلية ، فإن عقود الحلقة المتقلصة تؤدي إلى تكوين ثلم.

2. توسيع نظرية السطح:
اقترح ميتشيسون (1922) أن المواد النووية تحررها الصبغيات المسؤولة عن التوسع الخلوي في القطبين. مع توسع المناطق القطبية ، يتعاقد خط الاستواء الذي يؤدي إلى ظهور ثلم.
يقسم الأخدود الخلية إلى خليتين ابنتين. هناك بعض الأمثلة ، التي تبين أن تشكيل الأخدود يحدث في غياب النوى أو الكروموسومات (Nachtwey ، 1965). يشير هذا إلى أن مادة ماترينية مثيرة قد نشأت من غير النواة.

اقترح هذه النظرية دان ودان (1947) ودان (195 ب). وفقا لهم ، المغزل وأستريز هي المسؤولة عن السيتوبلازمية. أثناء التجربة ، كانت جزيئات الكاولين مرتبطة بغشاء البيضة.
لوحظ أن استطالة الخلية في الطور المتأخر مصحوبة بانكماش في المستوى الاستوائي ينتج جسيمتين من الكاولين على جانبي خط الاستواء يقتربان من بعضهما البعض. ويعتقد أن القوة الدافعة هي استطالة الأنابيب الدقيقة للمغزل مما يؤدي إلى تفريق المراكز.
4. نظرية الاسترخاء بالنجوم:
اقترح ووليبرت (1960 ، 1963) أن يكون سطح النداء تحت توتر منتظم. أثناء الانقسام الخلوي ، عندما تصل الأشعة النجمية إلى القطبين ، تنخفض في التوتر السطحي عند القطبين. يبقى التوتر السطحي عند خط الاستواء كما هو. بسبب التوتر السطحي المنخفض ، تتوسع المناطق القطبية وتسبب ظهور ثلم عند خط الاستواء.
5. نظرية تكوين الحويصلة:
وفقا ل Threadgold (1968) ، أثناء الطور ، تقسم خلية الانقسام. يحدث هذا في المنطقة بين المناطق بسبب زيادة كثافة الإلكترونات. كما يظهر غشاء البلازما كثافة إلكترون عالية في ثلم في نفس الوقت ، فإن ألياف المغزل المستمرة الموجودة في المنطقة المشتركة بين المناطق ، تستمر في زيادة كثافتها وتشكل في النهاية لوحة ليفية أديترونيك في خط الاستواء.
مع تقدم الأخاديد ، تظهر حويصلة غشاء كبيرة فارغة على كل جانب من هذه اللوحة. تضاف في وقت لاحق على الحويصلات الصغيرة في الطائرة الاستوائية. في المرحلة النهائية ، يحدث الانصهار في جميع الحويصلات الكبيرة والصغيرة لتشكيل ثلم عميق ، تاركة الخلايا البنتية متصلة بواسطة عروس بين الخلايا. علاوة على ذلك ، ينشأ صف من الحويصلات الصغيرة التي تسبب الفصل النهائي لخليتين ابنتين.

دور Centriole خلال الانقسام :
يعمل المركز كمركز مركزي لتطور المغزل المركزي. ما إذا كان المركب الوسطي ينتج ألياف المغزل أو يعمل على توجيه تكوينها أو من ناحية أخرى هو سلبي تماما في هذه العملية ، لا يمكن الاستدلال عليها من جميع المعلومات الموجودة.
ووفقًا لكليفلاند (1957) ، يمكن التعرف أولاً على الألياف المغزليّة والألياف النجمية والكروموسوماتية في المنطقة القريبة من السيقان الوسطى. ثم يعمل الجهاز الوني المتطور تمامًا لفصل الكروماتيدات البينية نحو القطبين المختلفين.
فيما يتعلق بالانقسام الفتيلي ، اختتم كليفلاند نقطتين هامتين:
(1) الاستقلال المؤقت النسبي للجهاز الوني واللوني فيما يتعلق بالازدواج الظاهري والوظيفة ؛
(2) الاعتماد النهائي للخلية على سلامة هذين النظامين ووظيفتهما الموحدة.
مدة الانقسام:
الوقت اللازم للانقسام يختلف مع الأنواع والبيئة. درجة الحرارة والتغذية ، على وجه الخصوص ، من العوامل الهامة. قد يتم إكمال التسلسل الكامل للمراحل في 6 دقائق إلى عدة ساعات. عادة ما تستغرق الدورة الكاملة للانقسام الخلوي حوالي 18 ساعة ؛ حوالي 17 ساعة للمرحلة. مراحل مختلفة من الانقسام هي مختلفة المدة. Anaphase هو أقصر ، الطور الطوري و telophase الأكثر لفترات طويلة ، والطبقة الطورية من المدة المتوسطة.
الطاولة. مدة مراحل Mitotic (في دقائق)
مثال | درجة الحرارة درجة مئوية | الطور الأول | الطورية | Anaphas | الطور النهائي |
الفأر | 38 | 21 | 13 | 5 | 4 |
دجاج | 39 | 30-60 | 2-10 | 3-7 | 2-10 |
(خلايا Mesenchyme | |||||
في زراعة الأنسجة) | |||||
الضفدع (الخلايا الليفية | 20-24 | 32 | 20-29 | 6-11 | |
في الثقافة) | |||||
الجراد | 38 | 102 | 13 | 9 | 57 |
(Neuroblasts) | |||||
قنفذ البحر الجنين | 12 | 19 | 17 ' | 2 | 18 |
البصل (طرف الجذر) | 20 | 71 | 6.5 | 2.4 | 3.8 |
البازلاء (طرف الجذر) | 20 | 78 | 14.4 | 4.2 | 13.2 |
1. التوزيع المتساوي للكروموسومات:
السمة الأساسية للانقسام الفتيلي هي أن الكروموسومات موزعة بالتساوي بين خليتي ابنتهما. مع كل انقسام خلوي هناك تقسيم للكروموسومات. العدد الثابت للكروموسومات في جميع خلايا الجسم هو بسبب الانقسام الفتيلي.
2. نسبة السطح إلى الحجم:
يستعيد الانقسام نسبة سطحية الخلية للخلية. تحتوي الخلية الصغيرة على قدر أكبر من المساحة المتاحة بالنسبة للحجم مقارنة بالخلية الكبيرة. ومع زيادة حجم الخلية ، تصبح مساحة السطح المتاحة بالنسبة إلى الحجم الزائد أقل. من خلال التقسيم تصبح الخلية أصغر في الحجم ويتم استعادة نسبة حجم السطح.
3. مؤشر nucleoplasmic :
نمو كائن متعدد الخلايا يرجع إلى الانقسام الفتيلي. لا يمكن أن تنمو الخلية في الحجم إلى حد كبير دون إزعاج النسبة بين النواة والسيتوبلازم. بعد الوصول إلى حجم معين ، تنقسم الخلية لاستعادة مؤشر النيوكليوبلازمية. وهكذا يحدث النمو من خلال زيادة عدد الخلايا ، بدلاً من الزيادة في حجم الخلايا.
4. إصلاح:
يحدث إصلاح الجسم بسبب إضافة الخلايا عن طريق الانقسام الفتيلي. يتم باستمرار استبدال الخلايا الميتة من الطبقة العليا من البشرة ، وخلايا بطانة القناة الهضمية ، وكريات الدم الحمراء. يقدر أنه في جسم الإنسان يتم فقدان حوالي 500،000،000،000 خلية يوميًا.