الكروموسومات: مورفولوجيا ، هيكل ، Heteropycnosis وتفاصيل أخرى
الكروموسومات: مورفولوجيا ، بنية ، مغاير ، صِبْغِيّ كروموسوم وبنية تحتية للكروموسوم!
شوهدت الكروموسومات لأول مرة من قبل هوفميستر (1848) في الخلايا الأم لحبوب اللقاح في Tradescantia على شكل أجسام ملطخة بدقّة. تم استخدام كروموسوم المصطلح (Gr: chrom = color؛ soma = body) بواسطة Waldeyer (1888) لتعيين انجذابهم الكبير للأصباغ الأساسية.
تم وصف أهميتها الوظيفية من قبل IV.S. ساتون (1900) عندما تتبع التوازي بين فصل الكروموسومات أثناء الانقسام الاختزالي وانتقال العوامل الوراثية خلال النشوء الجرمي. تم نشر مراجعات عامة حول مورفولوجيا الكروموسومات من قبل Heitz (1935) ، Kuwada (1939) ، Geitler (1940) و Kaufmann (1948).
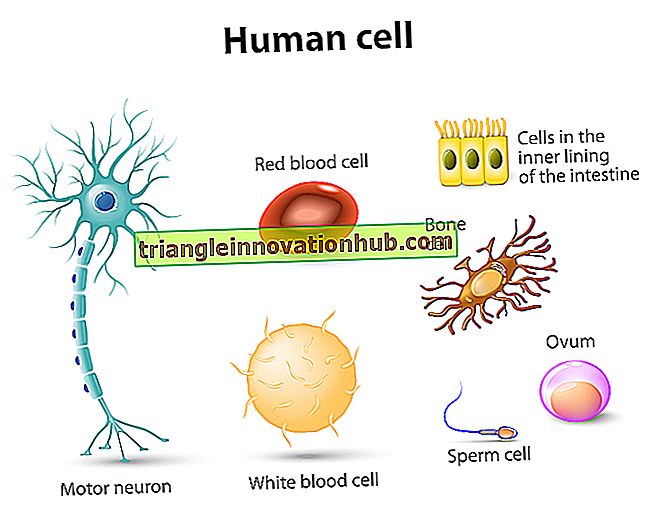
الكروموسومات هي أهم مكونات الخلية ، خاصة أنها تظهر أثناء الانقسام والانقسام الاختزالي. تم إثبات وجودهم قبل فترة طويلة من تسمية "فالديير" في عام 1888 باسم "الكروموسومات".
يمكن اعتبار الكروموسوم كعنصر نووي له تنظيم خاص وفردية ووظيفية. وهي قادرة على التكاثر الذاتي مع الحفاظ على خصائصها المورفولوجية والفسيولوجية من خلال التقسيمات الخلوية المتعاقبة.
علم التشكل المورفولوجيا:
يمكن أن يكون شكل مورفولوجيا الكروموسوم أفضل الدراسات عند الطورية أو مرحلة الطور من الانقسام الفتيلي عند وجودها كعضيات محددة ، كونها أكثر كثافة أو طليعة.
رقم :
عادة ما يكون عدد الكروموسومات في نوع معين ثابتًا يحتوي على عدد مضاعف (2n) من الكروموسومات في خلايا الجسم الجسدية ، وعدد الكروموسومات (النانوية أو المخفّضة) عدد (n) من الكروموسومات في خلاياها الجنسية (الحيوانات المنوية والبويضات). عدد الكروموسومات متغير من واحد إلى عدة مئات بين الأنواع المختلفة
على سبيل المثال ، في أسكارس megalocephala هو 2 ، بينما في بعض الأوليات (Aggreta) ، هناك أكثر من 300 كروموسوم ، في Paramecium 30 إلى 40 ، في radiolarians ما يصل إلى 1600 ، في Hydra vulgaris 32 ، Musca domestica 12، Rana esculenta 26 ، كولومبا ليفيا 80 ، cuniculus Oryctolagus 44 ، Gorilla gorilla 48 و Homo sapiens (رجل) 46.
تساعد أرقام الصبغي أيضًا على التصنيف. في كاسيات البذور ، أكثر عدد أحادي الصبغة هو 12 وعدد أعضاء هذه المجموعة تتراوح من 3 إلى 16. وبالمثل ، في عدد الفطريات ، يتراوح العدد الفرداني من 3 إلى 8.
في الرئيسيات ، يتراوح عدد الصبغة الفرداني من 16 إلى 30. هذه المجموعة الصبغية من الكروموسومات موجودة في نواة الأمشاج ، بينما في خلية ثنائية الصيغة الصبغية سيكون هناك جينومان. الخلايا ثنائية الصبغية هي الخلايا الجسدية في الجسم. تحصل الخلايا ثنائية الصيغة الصبغية على مجموعة الكروموسومات ثنائية الصبغة عن طريق اتحاد الأمشاج أحادي الصبغة من الذكور والإناث في التكاثر الجنسي.


بحجم:
تتراوح الكروموسومات ، في المتوسط من 0.5 إلى حوالي 30 µ في الطول ومن 0.2 إلى Зµ في القطر. يختلف العدد النسبي للكروموسومات عمومًا في النواة ، ولكن في وقت واحد قد تكون جميع الكروموزومات في الخلية من نفس الحجم. الخلايا النباتية تمتلك عادة كروموسومات أكبر ثم خلايا حيوانية.
يحتوي التريليوم على كروموسومات قد تصل إلى طول 32 درجة في الطورية. عادةً ما يكون للنباتات أحادية الفلقة كروموسومات أكبر من الصبغيات التي تحتوي على عدد أكبر من الكروموسومات. بين الحيوانات ، الجنادب ، الصراصير ، mantids ، و newts و salamanders ديها كروموسومات كبيرة.
يمكن تحفيز التباين في حجم الصبغيات بواسطة عدد من العوامل البيئية:
1. الخلايا التي تنقسم عند درجة حرارة منخفضة تحتوي على كروموسومات أقصر وأضيق من تلك التي تنقسم عند درجات الحرارة المرتفعة.
2. Colchicine هو قلويد الذي يتداخل مع تشكيل المغزل وانقسام الخلايا. يميل إلى تقصير الكروموسومات.
3. الانقسام السريع والمتكرر يؤدي إلى صبغيات أصغر. يبدو أن معدل الانقسام الخلوي يتقدم بسرعة أكبر من تكوين مادة الكروماتين كما هو معتاد.
4. في النباتات ، أثرت كمية الفوسفات في الوسط الغذائي على حجم الصبغيات. التركيز العالي يعطي كروموسومات أكبر من تلك النباتات التي تعاني من نقص الفوسفات. بما أن الفوسفات هو جزء لا يتجزأ من جزيء الحمض النووي ، فإنه يبدو أن كمية الحمض النووي في الكروموسوم يمكن أن تتنوع لإعطاء تغييرات في الحجم.
شكل:
يتغير شكل الكروموسومات من مرحلة إلى أخرى في العملية المستمرة لنمو الخلية وتقسيم الخلية. في مرحلة الراحة أو مرحلة الطور البيني للخلية ، تحدث الصبغيات في شكل هياكل قابلة للإنحناء ، مرنة ، ملفوفة وقابلة للتقلص ، مثل خيوط الكروماتين.
في الطورية و الطور المتأخر ، تصبح الصبغيات سميكة وخيطية. كل كروموسوم يحتوي على منطقة واضحة ، تعرف باسم centromere أو kinetochore ، على طول طولها. يقسم السنترومير الصبغيات إلى جزأين ، كل جزء يسمى ذراع الكروموسوم.
يختلف مركز centromere من كروموسوم إلى كروموسوم ، ويوفر أشكالًا مختلفة للأشكال التالية:
1. عبر مركز الاتصالات :
وتعرف الكروموزومات التي تشبه القضبان والتي لها السنترومير على الطرف القريب باسم الكروموسومات التي يتم إرسالها عبر الإنترنت.
2. Acrocentric :
فالكروموسومات الشيطانية الشكل تشبه J ، ولكنها تحتوي على المركزية في نهاية واحدة ، وبالتالي تعطي ذراعًا قصيرًا وذراعًا طويلة بشكل استثنائي. الجراد (Acrididae) لديه الكروموسومات acrocentric.

3. sub-metacentric :
تكون الصبغيات الفرعية الفرعية على شكل حرف L. في هذه ، يحدث centromere بالقرب من المركز أو في الجزء المتوسط من الكروموسوم وبالتالي تشكيل اثنين من الأسلحة غير المتكافئة.
4. metacentric :
تكون الكروموسومات المتقابلة في شكل V وفي هذه الكروموسومات يحدث المركز في الوسط ويشكل ذراعين متساويين. البرمائيات لها كروموسومات متقاطعة.
هيكل الكروموسوم:
في وقت سابق وصف أوصاف مجهرية كان يعتقد الكروموسوم ، أو الكروماتيد ، تتكون من خيط ملفوف دعا الكرومونيما الكذب في المصفوفة. كان من المفترض أن يكون الكروموسوم مشمولًا بأغشية pellicle غشائية.

في وقت لاحق أظهرت الدراسات المجهرية الإلكترونية أنه لا يوجد قاطع غشائي محدد يحيط بالكروموسوم. تشمل البنى الأخرى الموجودة في الكروموسومات ، الكروماتيدات ، الكادميومير ، التضيقات الثانوية ، المنظِّمات النووية ، التيلوميرات والسواتل على النحو الوارد تحت الرؤوس التالية:
الكروماتيدات :
خلال الطور التصاعدي يبدو أن الكروموسوم يمتلك خيوطين تدعى الكروماتيدات chromatids ، والتي تصبح متشابكة في مصفوفة الكروموسوم. يتم تثبيت هذين الصبغيين عند نقطة على طولهما في منطقة انقباض الكروموسوم.
هذه الكروماتيدات هي حقا chromonemata اللولبية coild (الغناء. ، chromonema) في الطورية. كان أول من لاحظ الشعيرات الملفوفة من قبل Baranetzky ، في عام 1880 ، في الخلايا الأم لحبوب اللقاح في Tradescantia وكان يطلق عليه chromonema بواسطة Vejdovsky في عام 1912.
يمكن أن تتكون الصبغيات من 2 أو 4 ليفات أو أكثر اعتمادا على النوع. هذا العدد من الليفونات في الكرومونيما قد يعتمد على المراحل المختلفة لأنه في مرحلة واحدة قد يحتوي على ليفل واحد و طور آخر قد يحتوي على اثنين أو أربعة ليفات. هذه اللفائف من chromonema ملفوفة مع بعضها البعض.
الملفات هي من نوعين:

1. ملفات paranemic :
عندما يمكن فصل الألياف الليفية عن بعضها بسهولة ، تسمى هذه اللفائف ملفات اللف.
2. لفائف plectonemic:
هنا تتداخل الألياف الليفية الكرومومية بشكل وثيق ولا يمكن فصلها بسهولة. تسمى هذه الملفات لفائف plectonemic. تعتمد درجة تجلط الالياف chromonemal خلال انقسام الخلايا على طول الكروموسوم.
هناك ثلاثة أنواع من اللفات:
(ط) تمتلك الملفات الرئيسية لل chromonema 10-30 جاما.
(2) تكون الملفات الصغرى لل chromonema متعامدة مع الملفات الرئيسية ولها العديد من الجيروسات كما هو ملاحظ في الصبغيات الميتة. إذا لم يحدث الانشقاق حتى الآن في هذه المرحلة ، سيكون هناك كروموسما واحد ، إذا كان قد حدث بالفعل سيكون هناك نوعان من chromonemata.
(3) يتم العثور على ملفات قياسية أو جسدية في الكرومونوما للانقسام الفتيلي حيث تمتلك chromonemata هياكل حلزونية تشبه الملفات الرئيسية للكروموسوم الانتصافي.
Chromomeres:
تم العثور على الكرومونيما للكروموسومات الرقيقة من الطور الانقسامي والميوزي أن تحتوي على مناطق سميكة ورقيقة بالتناوب ، مما يعطي مظهر قلادة فيه عدة حبات تحدث على الخيط.
تُعرف البنى السميكة أو الحبيبية للكرومونيوم بالكروموميرات ، وتسمى المناطق الرقيقة بين الكروموسيرات بالكروموميرات. تم العثور على موقف من الكروموسيرات في chromonema أن تكون ثابتة لكروموسوم معين.
وقد أعطى علماء الخلايا تجارب مختلفة حول الكروموسيرات. ينظر البعض إلى الكروموسيرات كمواد بروتين نووي مكثف ، في حين افترض البعض الآخر أن الكروموسيرات هي مناطق لفائف فائقة.
تم تأكيد الرؤية اللاحقة من خلال الملاحظات المجهرية الإلكترونية. لفترة طويلة ، اعتبر معظم علماء الوراثة هذه الكروموسومات بمثابة جينات ، أي وحدات الوراثة.
سنترومير :
وهو جزء لا غنى عنه من الكروموسوم ويشكل التضيق الأساسي في الطورية. بدون الصبغيات لا تستطيع الكروموسومات توجيه نفسها بشكل صحيح على اللوحة الطورية. وحيث أن الكادميومرات تحتل موقعًا ثابتًا ، فإن الكواكب تكون مسؤولة عن شكل الكروموسومات.
وهكذا ، يتم تحديد شكل الكروموسومات بواسطة التضيق الأساسي ، الذي يقع عند نقطة التقاء أذرع الكروموسومات. في داخل التضيق الأساسي ، هناك منطقة واضحة ، لها حبيبة صغيرة أو كروية. تعرف هذه المنطقة الصافية باسم centromere (Gr. meros، part) أو kinetochore أو kinetomere.
وظيفتها هي من حيث الحركة. وهي مسؤولة عن تكوين ألياف الكروموسومات في المغزل. هيكل السنتروم هو بيضاوي ، غير قابل للرسم ، ذو قطر كبير ، كما هو الحال في الذرة أو قد يكون مثل حبيبات صغيرة أو كروية ، كما هو الحال في Tradescantia.
في وسط السنتروم ، قد يكون هناك حبيبات أو كروية صغيرة أو أكثر ، تسمى الكروموسومات وألياف المغزل. عادة ، كل كروموسوم لديه centromere واحد فقط. في مثل هذه الحالات ، يسمى الكروموسوم أحادي القطب. قد يكون هناك اثنين ، أي ثنائي المركز أو أكثر متعدد المراكز أو مع centromere منتشر ، كما هو موضح في الصهريج الكبير في Ascaris و Hemiptera.
بعد الدراسات الحديثة ، كان من المعروف أن centromere يتكون من ثلاث مناطق موجودة في نسختين. تحافظ المنطقة الوسطى على علاقة الكروموسومات بالمغزل. يوضح الرسم التوضيحي أدناه وجود شقيقتين- chromatids يشكلان كل كروموسوم طوري يحمله إقليم له دورة تقسيم خاصة.

يعتبر السنترومير مقسوما وظيفيا إلى المحور الطولي للكروموسوم في بداية الطور. يحكم تحركها نحو الأقطاب من خلال تعلقها بالمغزل. في وقت ما ، تحدث الانقسامات أيضا في الزاوية اليمنى للمحور الطولي الذي يشكل اثنين من الكواكب ، التي تعلق فيها الكروماتين من كل ذراع.
هذه البنية ، المكونة من ذراعين ، تُعرف بالكروموسوم. واقترح اسم Darlingtion في عام 1939. Mc. وقد ذكر Clintock (1932) أن مثل هذا الكسر ممكن أيضا بالأشعة السينية. في مثل هذه الحالات ، كل جزء جزء من centromere وظيفية.
ومن المعروف أيضا أن السنترومير هو مركب مركب ، يتم تنسيق أجزاءه في التقسيم والحركة. اقترح Sachrader (1936) و Darlington (1939) أن centromere يمكن اعتباره متماثلاً مع centrioles على الهيكلية وكذلك على أساس النظرية.
الانقباض الثانوي :
بالإضافة إلى التضيق الأساسي أو centromere قد تظهر ذراعي الكروموسوم واحد أو أكثر من التضيقات الثانوية (تسمى ثانوي consisriction-II). هذه تختلف عن المنظمين النوويين (يسمى الانقباض الثانوي الأول) ، على الرغم من أن بعض علماء الخلايا يشيرون أيضًا إلى المنظم النووي باعتباره الانقباض الثانوي.
يكون موقع التضيق الثانوي الثاني ثابتًا لكروموسوم معين ، وهو بالتالي مفيد في تحديد الكروموسومات. وقد اقترح أن القيود الثانوية تمثل مواقع الكسر والانصهار اللاحق. في الرجل ، توجد ثغرات ثانوية على الأذرع الطويلة للكروموسومات 1 و 10 و 13 و 16 و Y منظم نوكليولار (انقباض ثانوي 1).
المنظم النوكليول (المرحلة الثانوية I):
عادة في كل مجموعة من الكروموسومات الثنائية الصبغيات ، يكون للكروموسومات المتجانسة "تقلصات" إضافية تسمى المنظومات النووية. هذه هي ما يسمى لأنها ضرورية لتشكيل النواة.
تتشكل النواة في مرحلة إعادة البناء بعد الإنقسام الفنتيكي. تحت المجهر الضوئي يظهر المنظم النوكليوليار على شكل "انقباض" بالقرب من طرف واحد من الكروموسوم. جزء من الكروموسوم خارج المنظم النووي قصير جدا ويظهر مثل كرة (قمر صناعي). في الكروموسومات البشرية 13 و 14 و 15 و 21 22 و Y لها منظومات نيوكليولية وأقمار صناعية. تسمى الكروموسومات التي تحمل الأقمار الصناعية SA كروموسومات T.
البادئة SAT تعني "Sine Acid Thymonucleionico" (بدون حمض ثيموكليونيك أو DNA) ، حيث أن الكروموسوم الموجود على التلوين يظهر عوزًا نسبيًا للحمض النووي في المنطقة المنظِّمة للنواة. هناك على الأقل اثنين من الكروموسومات SAT في كل نواة ثنائي الصبغة.
التيلوميرات :
نهايات الصبغي تتصرف بشكل مختلف عن الأجزاء الخلاليّة. إذا تم كسر نهاية أو تيلومير إما تلقائيا أو عن طريق الحث ، فإنه عادة ما يتم فقده من النواة في انقسام الخلية لاحقا لأنه يفتقر إلى المركزية.
تكون النهاية المكسورة للكروموسوم الحر المتبقي مستقرة وقد تتحد مع نهاية مكسورة أخرى للكروموسوم في المنطقة المجاورة. ومع ذلك ، فإن نهاية كسر لن وحدة مع نهاية عادية. في الطور الانفعالي ، تنجذب التيلوميرات أحيانًا إلى المركز وتنتقل إلى الغشاء النووي بالقرب من المركز. ينتج عن هذا السلوك ما تم وصفه بأنه مرحلة Bouquet.
ماريكس الكروموسوم :
كما يفترض من قبل بعض علماء الخلايا أن chromonemata مضمنة في مصفوفة لونية يحدها pellicle. ومع ذلك ، كشفت الملاحظة الأخيرة من الدراسات الميكروسكوبية الإلكترونية عدم وجود pellicle. لم يتم تعريف المصفوفة بأنها الكتلة الرئيسية للكروموسوم ، والتي هي مميزة بشكل إيجابي. يمكن إزالته بوسائل إنزيمية تاركا وراءه كروموسوم متبقي سلبي من Feulgen.
Heteropycnosis:
وقد لوحظ بشكل عام خلال مراحل مختلفة من الانقسام الفتيلي أن كروموسومات معينة أو أجزاء من الكروموسومات ليست كذلك ، ولكنها أكثر كثافة من بقية النمط النووي. هذا يشير إلى heteropycnosis. هذه الظاهرة تؤدي إلى تكتل الكروموسومات أثناء انقسام الخلية. قد يكون Heteropycnosis موجباً ، متبوعاً بالتكثيف أو السلبي ، مع إظهار أو عدم وجود تكاثف.
وقد لوحظ أيضًا أن جزءًا معينًا من الكروموسوم أو الكروموسوم بأكمله قد لا يظهر التكاثف أو التغاير في جميع المراحل. وبما أن تغيّر الحلقات المغايرة هو خصوصية للغيوم المتغايرة ، فإنه يساعد في ترسيمها من اليوكروماتين. إنه أكثر انتشارا في كروموسوم الجنس ، على الرغم من أن الآخرين يعرضونه أيضا.
يوتروماتين وهرموكروماتين :
على الرغم من أنه ، خلال المرحلة البينية ، ينتشر لونين من الكروموسومات في شكل خيوط رفيعة من الكتان ولكن في مناطق معينة ، تعرف الكروماتين بمناطق heterochromatin أو heterochromatin.
Heterochromatin هو من نوعين:
1. heterochromatin الاختياري ، و
2. heterochromatin التأسيسية.
1. heterochromatin الاختياري :
وهذا يمثل حالة مؤقتة من تعطيل الكروماتين الذي يصبح فيه كروموسوم واحد من الزوج مغايرًا جزئيًا أو كليًا. على سبيل المثال ، في الثدييات واحد من اثنين من الكروموسومات X في الخلايا الجسدية الأنثوية يصبح heterochromatic وتشكل نوع الجنس-جيني أو بار (بار وبرترام ، 1944). في الخلية الجسدية الذكرية ، هناك كروموسوم X واحد فقط ويبقى متشابهًا (بدون جسد بار).
2. heterochromatin التأسيسية:
هذا النوع من heterochromatin يقدم ميزة دائمة أكثر ويوجد في كل من الكروموزومات للزوج. غالباً ما توجد في المناطق القطبية الوسطى ، التيلوميرات ، في مناطق المنظِمات النووية أو كحزام في مناطق أخرى من الكروموسومات. يرتبط ارتباطا وثيقا مع نواة في كل من النباتات والحيوانات.
الكروموسوم النطاقات:
أدخلت TC Hsu وغيرها (1969) طرقًا جديدة لصبغ الكروموسومات التي أصبحت بها الأنماط المتميزة من العصابات الملطخة والخطوط البينية الملطخة قليلاً. كانت طرق التلوين هذه بالغة الأهمية لأنها سمحت بتحديد كل كروموسوم بشكل فريد ، حتى لو كان التشكل الكلي متطابقًا. يمكن الآن إجراء التمييز بين كروموسومات المجموعة الثانية المماثلة نسبيًا. على سبيل المثال ، قد نقول الآن أنا أو 2 أو 3 كروموسومات بدلاً من كروموسوم من مجموعة دينز.
ما يلي هو طرق الربط بالكروموسوم:
1. G-Banding :
إن طريقة ربط الصبغي الأكثر فائدة هي G-banding. تم تطوير هذه التقنية من قبل هسو وأريغي. لوحظ أنه عندما يتم تحضين الكروموسومات في اللعاب تكون ملطخة ببقع Giemsa أو معالجتها باليوريا أو المنظفات. تظهر G-Bands في المناطق التي هي بروتين S الغنية. الاستعدادات Giemsa ملطخة هي أكثر دائمة وتتطلب البصريات المجهرية العادية والإضاءة.
2. Q-Banding :
تم تطوير هذه التقنية من قبل Casperson. ويلاحظ ، عندما يتم صبغ الكروموسومات مع الخردل quinacrine وملاحظتها من خلال المجهر مضان ، تلوث مناطق الكروموسومات الغنية في الأدينين والثايمين بشكل مكثف.
مناطق الغوانين-السيتوزين لا تزال غير ملوثة. تسمى هذه المناطق Q-bands. عيب هذا التلوين هو أن البقع تتلاشى بعد فترة قصيرة ، وعلاوة على ذلك ، هناك حاجة إلى بصريات مجهرية خاصة بالإضافة إلى إضاءة فوق بنفسجية لرؤية هذه العصابات.

3. النطاقات:
تم تطوير هذه التقنية من قبل Pardue و Gall. يتم التعامل مع الكروموسومات مع هيدروكسيد الصوديوم قوية تليها المياه المالحة الدافئة ومن ثم ملطخة مع صبغة Giemsa. تتجلى هذه النغمات بشكل خاص حول السنترومير وفي الكروموسومات الأخرى التي تحتوي على كميات بديلة من الهيتروكروماتين التأسيسي المتكرر.
4. R. النطاقات:
تظهر هذه العصابات عندما يتم تحضين الكروموسومات في مخزن مؤقت عند درجة حرارة عالية وملطخة ببقعة Giemsa. تتوافق النطاقات R-band مع المناطق على charom9osomes التي تحتوي على بروتينات تفتقر إلى الكبريت. هذه هي المعاملة بالمثل من G العصابات.

تقنية الانحناء من الصبغ الصبغي مفيدة للغاية في معرفة أنواع مختلفة من الانحرافات الصبغية مثل الحذف ، الازدواجية ، الانقلاب أو النقل. وكثيراً ما يسمح اليقين الأكبر في تحديد الكروموسومات الكاملة أو أجزاء من الكروموسومات بواسطة G-bands للمحقق أن يعرف بالضبط أي الكروموسومات موجودة وأي أجزاء الكروموسوم خضعت لإعادة ترتيب هيكلية. يوفر التطويق أيضًا وسيلة لمقارنة الأنواع المتنوعة من الأنواع ذات الصلة ووصف الاختلافات التي يبدو أن لها أساس تطوري.
بنية فائقة للكروموسوم:
تم اقتراح وجهين للهيكل الفائق للكروموسومات:
(أ) عرض متعدد النقاط :
هذا ما اقترحه ريس (1966). بواسطة المجهر الإلكتروني ، أصغر وحدة مرئية للكروموسوم هي الفيبرال الذي يبلغ سمكه 100 درجة مئوية. يحتوي هذا الفيفيل على جزيئتي دنا مزدوجتين حلزونيتين يفصل بينهما فاصل 25A ° والبروتين المصاحب.
ثاني أكبر وحدة هي نصف الكروماتيد. يتكون نصف الكروماتيد من أربعة ليفات 100 A 0 بحيث يكون سمك 400 A ويحتوي على ثمانية حلزون مزدوج على DNA والبروتين المصاحب. اثنان نصف الكروماتيدات من الكروماتيد الكامل يتكون من 16 جزيء مزدوج من دنا الحلزون.
كما يتكون الكروموسوم من اثنين من الكروماتيدات ، وبالتالي سيكون العدد الإجمالي للحلزون 32 و dimeter 1600 A ° سميكة قبل الازدواجية أو التوليف. بعد كروموسوم الازدواجية لديه 64 الحلزون المزدوج من الحمض النووي مع قطر 3200 A المقابلة. يختلف عدد حلزون د ن أ في كل وحدة فوق مستوى الليفي باختلاف الأنواع. باختصار ، يتألف الكروموسوم من العديد من الألياف الدقيقة ، أصغرها هو جزيء بروتين نووي وحيد.



(ب) نموذج مطوي- Fibril:
قدم DuPraw (1965) هذا النموذج للبنية الدقيقة للكروموسمي. وفقا لهذا النموذج ، يتكون الكروموسوم من سلسلة طويلة واحدة من الحمض النووي والبروتين تشكيل ما يسمى ب fibril. يُطوى الليفرين عدة مرات ويؤثر بشكل غير منتظم على شكل الكروماتيد. هذا المقياس 250-300 A في السمك.
الوحدة الفرعية Nucleosome من الكروماتين:
أبلغ لونين من وحدات التكرار ، ودعا nucleosomes. وقد أعطى هذا المصطلح من قبل Oudet وآخرون ، (1975). يتكون النيوكليوسوم من الحمض النووي وبروتينات الهيستون. تشكل البروتينات جسيمًا أساسيًا وهو جزيئان من كل من بروتينات هيستون الأربعة. H2a H2b و H3 و H4. يحيط سطح الجسيم الأساسي بـ 1.75 دورة من الحمض النووي (200 زوج من القواعد).
يسمى DNA الذي يربط الجسيم الأساسي بحمض الوصلة. بروتين هيستون آخر ، يحد HI إلى الحمض النووي الرابط. (كورنبرغ وتوماس ، 1974). يقيس الجسيم الأساسي 40 ° A وقطر 80 A °. يقيس كامل النيوكليوسوم 55 درجة مئوية في الارتفاع و 110 A 0 في القطر.
الكروموسومات المتعددة
كان Balbiani في عام 1881 أول من لاحظ كروموسومات الغدد اللعابية في الغدد اللعابية من يرقة Chironomus. يقتصر هذا النوع من الكروموسومات العملاقة على نوع معين من الأنسجة الجسدية في الحشرات التي تنتمي إلى نظام Diptera.
عادةً ما يصلون إلى أكبر حجم لهم في النوى الكروية للغدة اللعابية اليرقية ، ولكن توجد نوى مشابهة في كثير من الأحيان في أنسجة أخرى مثل خلايا البطانة في الأمعاء ومشتقاتها ، والنبيبات المالطية وكذلك في العضلات والخلايا الدهنية الخ. مصطلح "الكروم المتعدد" من كولر هو أكثر تفضيلا من المصطلح العام لكروموسومات الغدد اللعابية.
بناء:
هيكل كروموسوم الغدة اللعابية له أهمية وراثية خلوية عظيمة. على طول كامل الصبغي هناك سلسلة من العصابات الداكنة بالتناوب إلى مناطق أخرى واضحة تسمى interbands. العصابات المظلمة تصبغ بشكل مكثف وتكون فولجن إيجابية. علاوة على ذلك ، تمتص الأشعة فوق البنفسجية عند 600 درجة مئوية. يمكن اعتبار هذه العصابات كأقراص ، لأنها تحتل القطر الكامل للكروموسوم.
هم من حجم متفاوت. العصابات الأطول لديها بنية أكثر تعقيدًا. وغالباً ما يشكلون ثنائيات ، وهما شريطان يقعان بجانب بعضهما البعض ولسماكة وشكل متماثلين. و interbands هي من الجانب fibrillar ، لا تلطيخ مع الأصباغ الأساسية ، هي Feulgen سلبية وامتصاص القليل جدا من الضوء فوق البنفسجي. علاوة على ذلك ، فإنها توفر مرونة أكبر من مناطق النطاقات. وثابتة في حالة وتوزيع الأقراص أو العصابات في اثنين من الكروموسومات المتماثلة (المقترنة) أمر ملحوظ.

في حالة الدروسوفيلا ميلانوجاستر ، تظهر الصبغيات في كل نواة متعددة الأضلاع ، عندما يتم تسطيحها ، على شكل خيوط طويلة رفيعة وواحد قصير جداً متصلاً بكتلة مركزية تُعرف باسم مركز الكروموسوم ، والتي ترتبط بها أيضًا نواة كبيرة واحدة. العلاقة بين هذه الخيوط هي الكروموسومات الخفيفة من المجموعة الانقسامية العادية لهذه النتوءات ليست واضحة في البداية.
يعتمد التفسير على حقيقتين:
(1) يتم دمج العضوين في كل زوج من الكروموسومات عن كثب طوال طولهما ؛
(2) يتم دمج كل الكواكب الصبغية في جميع الكروموسومات مع الشرائح المتغايرة المجاورة لها لتكوين مركز الكروموسومات.

وهكذا ، من بين السلاسل الستة ، يمثل المصطلح القصير الكروموسومين الرابع الصبغي ، والأطول يمثل الكروموسومات X ، بينما الأربعة المتبقية هي أطراف الكروموسومات الثانية والثالثة على شكل V. في نوى الغدد اللعابية من يرقات الإناث ، فإن الحبلا الذي يمثل "X" هو ضعف ، مثل الآخرين ، في حين أنه في النواة من الذكور يكون فردًا واحدًا. يكون V صغيرًا تمامًا ويتم تضمينه تقريبًا تقريبًا في مركز chromo.
يحدث مركز كرومو في جميع أنواع ذبابة الفاكهة وحجمها اعتمادًا على ما إذا كانت الأجزاء المتغايرة القريبة أو غير واسعة النطاق. في بعض المجموعات الأخرى من Diptera ، تغيب عن مركز Chiadoceridae للأسرة و Chironomidae.
Puffis and Balbiani حق وجين النشاط :
أهم الخصائص المورفولوجية للكروموسوم المتعدد الأضلاع هو وجود العصابات والأشرطة المتداخلة. وقد وجد برور وبافان وبيرمان وماكهيلك وآخرون أنه في مراحل معينة من تطور اليرقات ، تظهر بعض النطاقات المحددة من الكروموسوم متعدد الأضلاع تضخماً.
تعتبر هذه العصابات المتضخمة وحدات مطلقة للوراثة - الجينات في العمل. تأخذ هذه الجينات النشطة شكل نفث منتشرة هنا وهناك على طول كروموسومات الغدد اللعابية. وقد وجد Beermann و Clever (1964) أن النفخات تنتج الحمض النووي الريبي والرنا RNA في نفخة واحدة يختلف عن الحمض النووي الريبي من نفخة أخرى.
وقد أظهرت ملاحظات من نفث أنماط نشاط الجينات في العديد من الحشرات النامية. كما يلاحظ أن بعض الهرمونات والمواد الأخرى يمكن أن تبدأ وتوقف وتمنع بعض هذه الأنشطة. يمكن أن يختلف التركيب الدقيق للفرقة الفردية فيما يتعلق بالنفحات الموجودة في مكان واحد على كروموسوم في نسيج واحد وفي مكان آخر على نفس الكروموسوم في وقت آخر أو في نسيج آخر. وقد لوحظ هذا التعديل الموضعي في بنية الكروموسوم من مختلف Diptera منذ عدة سنوات في وقت سابق ولكن تم التغاضي عن أهميتها المحتملة.
يتم تخفيف تماسك خيوط الكروموسومات في المناطق المنفوخة. تبدأ الحلقة السائبة دائمًا في شريط واحد. في نفث صغيرة ، وفرق معينة ببساطة يفقد كفاف حاد ويعرض منتشر ، من مظهر التركيز في المجهر. في مواضع أخرى أو في أوقات أخرى قد تبدو الفرقة وكأنها "تعرضت" إلى حلقة كبيرة أو حلقة حول الكروموسومات.
تسمى هذه الجوز مثل الهياكل حلقات Balbiani ، بعد EG Balbiani ، الذي وصفها لأول مرة في عام 1881 ؛ يُعتقد أن النفخ يرجع إلى ظهور أو تفكيك كروموسوم فردي في شريط. عند ملاحظة أن الأنسجة المحددة ومراحل التطور تتميز بنمط نفخة واضح ، افترض Beermann (1952) أن تسلسل معين من النفخات يمثل نمطًا مطابقًا لنشاط اللعبة. في الواقع ، يحدث التنشيط الجيني التفاضلي ، يمكن للمرء أن يتنبأ بأن الجينات في نوع معين من الخلايا ستنتفخ بانتظام في حين أن الجين نفسه في الأنسجة الأخرى لن ينفخ.
تم وصف جين من نفس الطبيعة في مجموعة من أربع خلايا من الغدد اللعابية في Chironomus. تنتج Chironomus pallidivittatus إفراز حبيبي. الأنواع ذات الصلة وثيقة Chironomus tentatus يعطي قبالة واضحة ، وعدم إفراز حبيبات من نفس الخلايا.
في الهجينة من هذين النوعين هذه الطبيعة يتبع قوانين Mendelian بسيطة من الوراثة. تمكن Beermann و Clever (1964) من تحديد الفرق في مجموعة من أقل من 10 فرق في واحد من كروموسينات Chironomus وأن الكروموسوم تم تعيينه ككروموسوم IV.
تمتلك الخلايا المنتجة للحبيبات من C. pallidivittatus نفخة مرتبطة بهذه المجموعة من العصابات ، وهي نفخة غير موجودة تمامًا في الموقع المطابق للكروموسوم الرابع في Chironomus tentatus. في الهجينة لا يظهر النفخ إلا على الكروموسوم القادم من الوالد C pallidivittatus ؛ ينتج الهجين عددًا أصغر بكثير من الحبيبات عن الوالد.
علاوة على ذلك ، فإن حجم النفخ يرتبط ارتباطًا إيجابيًا بعدد الحبيبات. هذا يكشف بشكل واضح تماما العلاقة بين النفخة ومنتج خلوي (محدد). توضح هذه الدراسة علاقة محددة بين البافجين والوظيفة المحددة للخلية.
نظريات حول بنية الكروموسوم متعدد الخلايا:
هناك ثلاث نظريات لشرح بنية الكروموسوم متعدد الكروموسومات.
من بينها ، التفسير الثالث هو في الواقع مزيج من النظرين الأولين:
1. الكروموسومات المتجمعة هي نتيجة لدورة متعددة من التكاثر الكروموسومي داخل الخلايا وتتألف من حزم من الكروموسومات العادية المطوية. هذه هي نظرية الإحتجاجات المتعددة التي ترعاها Her-twig (1935) ، كوبر (1938) Painter (1939) و Beerrnann (1952).
2. الكروموسومات المتعددة الأشكال هي كروموسومات مقترنة ذات طول وعرض هائلين بإضافة أو دمج مواد إضافية غير موجودة في الكروموسومات العادية. هذا هو المفهوم السنخي السابق لميتز (193 5) واقترحه كوداني (1942) ودارلينجتون (1949).
3- تتكون الكروموزومات المتجمعة من حزم من الكرومونيما ، ويعزى حجمها جزئياً على الأقل إلى تراكم مواد إضافية في مركز الكروموسومات ، وإلى نمو فعلي في طول الصبغة الكروموسية (Koltzof، 1934؛ Painter، 1934؛ كالفن وآخرون ، 1940 ؛ ريس ودورة ، 1954 ؛ وايت ، 1945)
نظرية التعدد:
يعتقد الرسام (1941) أن الزيادة في القطر ترجع إلى تضخم ، وربما بسبب ازدواجية مستمرة في الكروموسومات الفردية كوخ فصلًا متغايرًا للكرومونيماتا الفردي. وبالتالي في سياق التطور ، يتم حل كل كروم كروم أصلي عن طريق الفصل من خلال التمدد إلى عدد من الكروموسومات الأصغر.
تنتج مضاعفة تكبير وتراكم الكروموسومات المتجانسة مظهر أشرطة لونية عرضية. وبالتالي ، يصبح الكروموسوم متضاعفًا كالبوليتونيوم ، ولكن الكرومونات الذاتية الفردية ، والتي قد تصل إلى ما يصل إلى 1024 ، وفقًا لرسامين Painter ، في حين قدر Beermann (1952) أن درجة polytene تصل إلى 16000 مرة.

فرشاة مصباح الكروموسومات:
خلال مجموعة الحيوانات الفقارية ، تظهر الكروموسومات الجسدية البنية المعتادة ، ولكن ضمن البويضات النامية لتلك الفقاريات التي تمتلك بيضة يولكية وخلال مرحلة الانقسام الديسيوسي ، تتعرض نفس الصبغيات إلى تغير ملحوظ ، خصوصًا خصائص الزيادة الهائلة في الطول والطول. ظهور الشعر يشع ؛ أو الحلقات التي يبدو أنها تنظم نفسها من مظهر كروموسى تشبه الفرشاة أثناء الطور المايوزى.
تم وصف هذا النوع من الكروموسوم لأول مرة بالفلم (1882) و Rukert (1892) يُطلق على الاسم الشهير ، 'Lampbrush ، Ris (1951) وجد كروموسومًا مشابهًا في أسماك القرش والطيور والبرمائيات وما إلى ذلك. في بعض الأحيان تصل هذه الكروموسومات إلى أقصى حجم يصل إلى 800 إلى 1000 µ لكل كروموسوم.

يمتلك كروموسوم الفرشاة مصباحًا محورًا كروموسوميًا مركزيًا يُطلق عليه سلسلة من الحلقات الجانبية. يبدو أن الحلقات الخروج من منطقة كثيفة. وفقا ل Duryee ، كل كروموسوم يشبه اسطوانة بلاستيكية واحدة والتي ، في مواضع محددة ، هي حبيبات الكروماتين المضمنة.
في الكروموسومات ثنائية التكافؤ يوجد حوالي 150-200 حبيبة مقترنة. هذه الحبيبات هي أحجام اثنين ، أي ، chromioles أصغر و chrormioles أكبر ، في وقت لاحق هي ellipsoidal في المظهر كما لو تم عصره داخل المصفوفة.

تعطي الحلقات الجانبية مظهرًا يشبه الفرشاة. تُعتبر الحلقات مادة لونين مركَّبة للاستخدام الخارجي ، وليست جزءًا لا يتجزأ من chromonemata الممتدة في شكل ملف رئيسي كما اقترحه ريس (1945).
تعتمد نظرية Duryee (1941) على فرضية التوليف الجانبي على حقيقة أن تمدد الكروموسوم بواسطة التلاعب الدقيق أو التقلصات بواسطة أيونات الكالسيوم لا يتسبب في اختفاء الحلقات أو التشرد وأن حل الحلقات بواسطة مجموعة متنوعة من المواد إلى لا تؤثر الحبيبات على سلامة chromonemata.
أظهرت غال (1956) بشكل قاطع من خلال الفحص المجهري الإلكتروني أن الحلقات هي أجزاء من الكرومونماتا وأن اختفاءها الظاهر يرجع إلى حقيقة أنه قبل الاتصال قاموا بإلقاء طلاءهم من الحمض النووي.
يمتلك الكروموسوم مرونة ملحوظة. الحلقات الجانبية الممتدة من الكروموسيرات أكثر هشاشة. وقد فسر غال (1958) أن تشكيل الحلقات هو تغير فيزيولوجي يمكن عكسه وهو على الأرجح غير جيني.
ومع ذلك ، فهو يشير إلى أن الحلقات تظهر اختلافًا في مورفولوجيتها مما يشير أيضًا ، من ناحية أخرى ، إلى أن كل زوج من حلقات الحلقة ، يمثل موضعًا جينيًا مختلفًا مسؤولًا عن تكوين بعض المنتجات الخلوية المعينة.
يحتوي الكروموسوم المصنوع من الفرشاة على محور رئيسي مركزي متواصل مرن. يحيط المحور الدائري بالبروتين مع RNA. ربما تتعامل الحلقات بشكل رئيسي مع توليف الرنا والبروتين ومواد الصفار.
ولتفسير الحجم الكبير لحلقات الفرشاة بالمرشحات ، افترض كل من Callan و Loved (1960) أن كل منهما لا يتكون من جين ، بل عدد من النسخ المتكررة ، مرتبة بشكل خطي ، لجين واحد. الكروموسومات ، الوحدات الأساسية للمنظمة في الكروموسومات فرشاة مصباح موجودة في شكلين. هناك نسخة "رئيسية" من جينة معينة في كرومومير تشبه نسخة الجينات "العبد" المتطابقة.
بهذه الطريقة تحتوي حلقة على عدد من النسخ المكررة فقط. يتم عزل الجينات بخط مزدوج وخطوط عرضية مفردة تشير إلى نهايات النسخ المكررة. لاحظ غال و Callan أن الحلقات الجانبية لها دائما نهاية واحدة رقيقة وأحمق أكثر بكثير عند نقطة الإدراج في الكروموميري.
ويعتقد أيضا أن الحلقة التي خرجت من الكروموميري توحيدها في النهاية السميكة ، مما يدل على تراكم كثيف للجيش الملكي النيبالي. اقترح كالان كذلك أن نسخ "الرقيق" فقط هي التي تشارك في تخليق الحمض النووي الريبي. هذا يضمن احتمالات لتجميع كمية كبيرة من حمض الريبونوكلييك.
يظهر تكوين النيوكليولي في كروموسوم لامب بروش نمطا غير عادي. قد يكون هناك عدة مئات من النيوكليولي العائمة حرة في النواة. لم يتم فهم المغزى بشكل جيد ، ولكن من المشكوك فيه أنه يجب أن يكون تجميع المواد اللازمة للنمو.

ملحق أو الكروموسومات الزائدة:
تمتلك نوى بعض الحيوانات والنباتات ، بالإضافة إلى الصبغيات الطبيعية ، واحد أو أكثر من الكروموسومات التبعية أو العددية. كان ويلسون (1905) أول علماء فسيولوجيا يراقبهم في حشرة الهيمبتيرا ، ميتابوديوس. ومنذ ذلك الحين ، تم الإبلاغ عنها في العديد من الحشرات وفي العديد من النباتات العالية أيضًا.
في بعض الحالات ، طبيعتها وأصلها معروفان بالتأكيد. ومع ذلك ، فإن أصولهم غير معروفة بالكامل. وعادة ما تكون الصبغيات الزائدة عن بعضها أصغر من الأحجام الموجودة في أنواعها. يُنظر إلى أنها تؤدي بعض الوظائف ، التي تقوضها حتى الآن ، وهي وظيفة بسيطة للغاية لا يمكن الكشف عنها جينيًا.