النواة: مورفولوجيا ، هيكل ، التركيب الكيميائي ، وظائف وأهمية النواة
النواة: مورفولوجيا ، هيكل ، التركيب الكيميائي ، وظائف وأهمية النواة!
الميزة الأبرز للخلية عندما ينظر إليها تحت المجهر هي النواة. في الأصل تم الكشف عنها من قبل Leeuwenhoek في 1700 كأجسام متراجعة في مركز كريات الدم من سمك السلمون.
هذه الهياكل ، التي يجب أن تكون نواة ، حيث ينظر إليها مع العدسات البسيطة التي تم العثور عليها في وقت فراغه. لاحظ فونتانا في عام 1781 أجسامًا بيضاوية مشابهة داخل خلايا جلد ثعبان السمك ، وقدم الوصف الكامل من قبل عالم النبات الاسكتلندي روبرت براون في عام 1835.
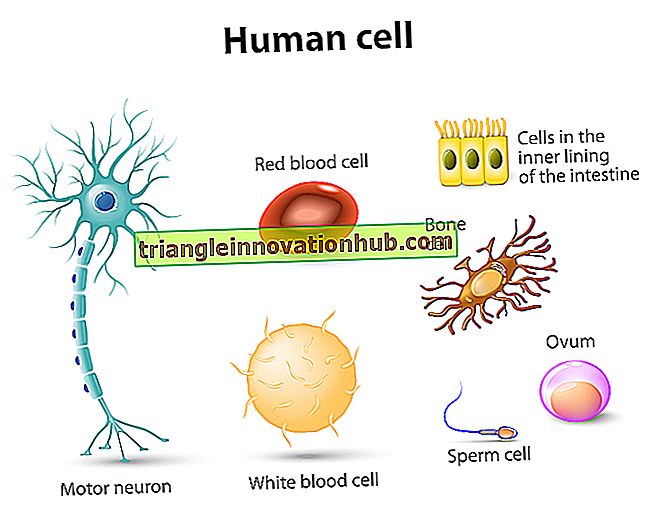
وهو عبارة عن بنية شبه عالمية للخلايا في وقت ما خلال دورة حياتها ، على الرغم من أن خلايا مثل الأنابيب الغربالية للنباتات الوعائية وخلايا الدم الحمراء للثدييات قد تفقد نواتها في الوقت الذي يتم تمييزها تمامًا.
النواة ليست متجانسة ، ولكنها تحتوي على خيط مثل الهياكل التي تم اكتشافها بعد بضع سنوات. يمكن الحصول على هذه التراكيب بصبغات الأنيلين التي تصبح معروفة بالكروموسومات (Gr-chroma = color و Soma = body). اقترح ساتون عام 1903 ، أولاً ، أن العوامل الوراثية أو الجينات تحمل على الكروموسومات.
وفي عام 1871 وللمرة الأولى تم عزله من قبل ميشر ، وكان تحليله الصرفي أقل تقدمًا بكثير من تحليل السيتوبلازم. تمكن <كسل> من إثبات وجود الحمض النووي ، ومنح هذا جائزة نوبل في عام 1910.
الخلية بدون النواة لها مستقبلات محدودة للغاية. النوع الوحيد من الخلايا الحيوانية الشائعة بدون نواة ، وهي خلايا الدم الحمراء في الثدييات ، يعيش فقط لمدة 3 إلى 4 أشهر. بغض النظر عن دورها في نقل الأوكسجين ، فهي مقيدة للغاية في أنشطتها الأيضية.
قد تنقسم خلايا البويضات التي أزيلت منها النوى تجريبيا لفترة من الزمن ، لكن منتجات التقسيم لا تفرق أبداً إلى أنواع خلايا متخصصة ، وفي النهاية تموت. الأجزاء التي لا تحتوي على نواة ، مقطوعة من كائنات كبيرة غير خلوية مثل الأميبات أو الطحالب ، Acetabularia ، تبقى حية مؤقتًا ، لكنها في النهاية تموت إلا إذا تم زرع نوى من خلايا أخرى فيها.
النواة ضرورية لاستمرار عملية الأيض على المدى الطويل وإلى قدرة الخلايا على تغيير هيكلها ووظائفها بشكل كبير (كما هو الحال في التمايز). في جزء كبير ، وهذا يعكس الدور الأساسي للنواة في إنتاج الحمض النووي الريبي المطلوب لتخليق البروتين.
عندما تتغير الخلايا ، فإن الوظائف والهياكل الجديدة تتطلب بروتينات جديدة. حتى الخلايا التي هي ثابتة في عملية التمثيل الغذائي والهيكل تظهر الاستبدال المستمر (دوران) الجزيئات الضخمة وربما العضيات ، بما في ذلك أجزاء من آلية تجميع البروتين السيتوبلازم.
علم التشكل المورفولوجيا:
شكل:
بشكل عام ، تكون النواة كروية ، ولكنها قد تكون مغزلية الشكل ، أو بيضاوية الشكل ، أو مسطحة ، حسب شكل الخلية ووظيفتها. في الخلايا الشابة ، غالباً ما تكون كروية ومركزية ، لكن تختلف عن بعضها البعض ، يمكن أن تكون مشرّدة وغير منتظمة الشكل.
قد تكون ممدودة النواة كما في ظهارة عمودي وخلايا العضلات. في بعض الحالات ، تصبح غير منتظمة وقد لوحظت في حالات معينة للتغيرات البطيئة في الشكل في الخلايا الحية ، على سبيل المثال ، خلايا الغضاريف ، الكريات البيض أو البويضات الحيوانية.
تكون نواة (Nucleus- singular) ذات شكل غير منتظم أو أموبي متكرر في الخلايا التي تتميز بعملية الأيض النشطة للغاية ، وفي هذه الحالة تكون النوى في الغالب ليس فقط ذات حجم كبير ، ولكنها تظهر زيادة إضافية ملحوظة في السطح عن طريق تكوين الفصوص ، التكات ، أو حتى في الحالات القصوى من الفروع المعقدة التي تتجسد في الخلية.
ويتم تقديم مثال متطرف على ذلك من خلال غدد يرقات معينة من الحشرات (Lepidoptera و Trichoptera) ، حيث تفترض النواة ، ذات الشكل الكروي الأصل ، أخيراً مظهر متشابك مع تشابكات تحتل مساحة كبيرة في الخلية.

في أنواع معينة من الخلايا ، يمكن أيضًا زيادة سطح النواة عن طريق تفتيته إلى حويصلات منفصلة أو حبيبات منفصلة أكثر أو أقل ، وبالتالي تكوين نواة "متعددة الأشكال" أو عش نووي. تحدث نوى حويصلية أو كروية الشكل بشكل عام في الخلايا النسيجية لمعظم الحيوانات والنباتات متعددة الخلايا. تحدث النوى الضخمة بشكل نموذجي في الخلايا الجنسية الذكرية للحيوانات عمومًا وفي العديد من النباتات المنخفضة (في نباتات coenocytic مثل Vaucharia وغيرها).
موضع:
يتم تحديد موقف النواة من خلال العديد من الأسباب ، مثل التوتر السطحي ، وموضع الفجوة ، والكثافة النسبية للسيتوبلازم في جزء مختلف من الخلية. في الخلايا الجنينية ، تحتل المركز الهندسي دائمًا تقريبًا.
في الخلايا غير المجوفة للنباتات حتى أنها تحتل مركز الكتلة السيتوبلازمية. في الخلايا الدهنية ، في البيض الغني بالصفار ، تُجبر النواة باتجاه المحيط عن طريق تراكم paraplasm. في الخلايا الغُدِّية ، يوجد في الوضع القاعدي ، الحبيبات تحتل السيتوبلازم القمي. يرتبط موضع النواة أيضًا بوظيفة الخلية.
بحجم:
في عام 1895 أظهر بوفيري أن حجم النوى في يرقات اليوشينيرم يعتمد على عدد الكروموسومات التي يحتويها كل منها. غير أن غيتس 1901 قدم أدلة تثبت أن هذه القاعدة ليست عالمية بأي حال من الأحوال. حجم النواة متغير ، ولكن بشكل عام يحمل بعض العلاقة مع السيتوبلازم. هذا يمكن التعبير عنه عدديا في ما يسمى مؤشر nucleoplasmic (NP) بواسطة O. Hertwigi 1906).
NP = Vn / Vc-Vn
حيث Vn يجري حجم النووية و Vc كونها حجم الخلية.
يوضح مؤشر NP هذا العلاقة بين حجم السيتوبلازم ونواة النواة ، أي إذا كانت الزيادات السابقة عندئذٍ ينبغي أن يزيد الثاني أيضًا. وتسمى نسبة حجم النواة إلى حجم السيتوبلازم في الخلية بـ "نسبة النيوكليوبلازمي" أو "نسبة الكربوبلازم".
رقم:
يعتمد عدد النوى الموجودة في أي كتلة من البروتوبلازم بشكل كبير على الجزء الأكبر من الكتلة حيث أنه يجب أن يتم الحفاظ على نسبة معينة من السطح النووي إلى الحجم السيتوبلازمي للعمل السليم للنظام بأكمله.
عموما جميع الخلايا هي uninucleate أو mononucleate أي ، نواة واحدة في خلية واحدة ولكن في حالات معينة تحدث حالة binucleate كما هو الحال في Paramecium caudatum ، وهنا نواة واحدة أصغر وهي النوى الصغيرة في حين أن أكبر واحد يسمى الكلي أو meganucleus. في حالات أخرى قليلة توجد حالة poly-or multinucleate كما في Opalina ، ألياف عضلية مخططة إلخ.
بناء:
تتكون النواة من أربعة مكونات:
(1) الغلاف النووي ،
(2) النسغ النووي ،
(3) Nucleolus ، و
(4) شبكة الكروماتين.
1. الغلاف النووي:
لم يتمتع الغلاف النووي إلا في الآونة الأخيرة بكثير من التحقيق. كما يتم عرضه مع مجهر تباين الطور القياسي ، فإنه يظهر أكثر من خط مظلم ، وهذا الخط الذي يفصل المحتويات النووية عن السيتوبلازم. في معظم الخلايا ، لوحظ أن تتحلل والإصلاح كما مراكز الخلايا وتكمل التقسيم. ومع ذلك ، في بعض الطحالب ، الأوليات والفطريات ، لا ينهار أثناء الانقسام.
تبين الملاحظة باستخدام المجهر الضوئي أن الغلاف النووي يعمل كحاجز بين النواة والسيتوبلازم. إن الطريقة التي تنظم فيها النواة تدفق المادة بدأت للتو في الدراسة. تكشف الدراسات الفائقة الهيكلية أن الغلاف النووي يتكون في الواقع من غشاءين منفصلين بفراغ محيط النواة يتراوح بين 110 و 400 درجة عرض.
يبدو أن الغشاء الداخلي على اتصال مع الكروماتين النووي ، الذي يمكن رؤيته مكثفًا على سطحه. الغشاء الخارجي مستمر مع الشبكية الهيوليوبلازمية السيتوبلازمية وغالبا ما يلاحظ مع الريبوسومات المرتبطة بسطحه.
ينتج عن ارتباط الغشاء الخارجي بالـ ER استمرارية بين النواة والسيتوبلازم. ومع ذلك ، فإن هذه الاستمرارية ليست كاملة بسبب وجود الغشاء النووي الداخلي ، الذي لا يزال بمثابة حاجز هيكلي.
المسام النووية :
وبما أن تخليق حمض الريبونوكلي يحدث داخل النواة بينما يحدث تخليق البروتين في السيتوبلازم ، فمن الواضح أن المواد يجب أن تكون قادرة على التحرك إلى الخارج. في الواقع ، هناك تدفق للمواد في كلا الاتجاهين من خلال المسام النووية ، والتي ، على الرغم من حجمها ، ليست قنوات مفتوحة على مصراعيها.
يتم دمج اثنين من الأغشية النووية معا على فترات لتشكيل المسام حوالي 600 ألف درجة مئوية في القطر. وقد شوهدت لأول مرة من قبل كالان وتوملين (1950). تشير التقديرات إلى أن المسام تشغل حوالي 10٪ من المساحة السطحية للغشاء النووي أو قد يختلف العدد الكلي للمسام في النواة من 100 إلى 5 × 10 7 . تظهر هذه المسام كمناطق دائرية أو متعددة الأضلاع.

ومع ذلك ، فإن المسام ليست مجرد ثقوب في الغشاء النووي. يشغل جزء كبير من الفتحة بترتيب أسطواني أو شبيه بالحلقة (الحلقي) للحبيبات ذات الكثافة المتوسطة أو المواد الليفية التي تبدو متواصلة مع الصفيحة الليفية الداخلية.
يتم تنظيم المواد الحلقيّة كمجمّع مسامي. تتكون الحلقة من ثماني حبيبات حلزونية على سطحها النووي والسيتوبلازمي وحبيبة مركزية في المركز. تمتد الألياف من الحبيبات المركزية والمواد الحلقية.
بعض المواد غير المتبلورة تشكل غشاء فوق المسام. أهمية الغشاء والحبيبات غير مؤكدة لأنها لا تبدو مكونات عالمية للمسام النووية. ربما تكون هياكل عابرة موجودة في بعض الحالات الفسيولوجية وليس في حالات أخرى.
ليفينا لامينا:
تعرض الأغشية النووية للعديد من الخلايا طبقة إضافية تسمى الصفيحة الليفية. إنه يعارض الجانب الداخلي للغشاء النووي الداخلي الذي يواجه المناطق الداخلية للنواة. وهو يتألف من خيوط دقيقة تتكون من البروتينات. على الأرجح أنها توفر التعزيز الميكانيكي للأغشية النووية.
وفقا لبعض العمال فإن الصفيحة الليفية تؤثر على تبادل المواد بين النواة والسيتوبلازم. درجة تطور الصفيحة الليفية تختلف اختلافا كبيرا في أنواع مختلفة من الخلايا. أما في خلايا الثدييات ، فهي رقيقة ولكنها عالية التطور في الأميبا وبعض اللافقاريات الأخرى. في Amoeba ، لديها تكوين يشبه قرص العسل وسماكة 1000 - 1500 A °.

نفاذية المظروف النووي :
تشير عدة تجارب إلى أن مجمعات المسام قد تكون فتحات مؤقتة أو دائمة في الغلاف النووي. عن طريق حقن جزيئات الذهب الغروية ، متفاوتة في الحجم من 2.5 إلى 17 نانومتر ، إلى سيتوبلازم الأميبا ، وجد أن تلك التي لديها أقطار تصل إلى 8.5 نانومتر دخلت بسرعة النواة.
توغلت الجسيمات بأقطار من 8.9 إلى 10.6 نانومتر ببطء أكثر ، ولم تدخل الأقطار الأكبر على الإطلاق. تشير هذه النتائج إلى أن الفتحات أصغر من حجم المسام. وقد تم الحصول على أدلة باستخدام هذه التقنيات ، مما يشير إلى أن المسام هو مسارات لتبادل الجزيئات الكبيرة. قد تنظم الزريعة التبادل بالنسبة للحجم وربما للطبيعة الكيميائية للمادة المخترقة.
من المهم اعتبار نفاذية الغلاف النووي غير ثابتة ، ولكنها تختلف باختلاف أنواع الخلايا وداخل خلية معينة على الأقل أثناء دورة الانقسام. وتعزى هذه الاختلافات إلى تغيرات في طبيعة المواد الحلقيّة (Feldher، 1971).
يجب أن يرتبط وجود المسام في الغلاف النووي ببعض من الخواص الكهروكيميائية لهذا الهيكل ، والتي يمكن بحثها عن طريق الخطوط الدقيقة.
تم التعرف على نوعين من المظاريف النووية بهذه التقنية. عندما يتم اختراق الخلايا العملاقة من الغدة اللعابية من ذبابة الفاكهة مع microelectrode ، هناك تغيير مفاجئ في إمكانات في غشاء البلازما (-12 فولت) ؛ ثم ، عندما يدخل microelectrode النواة ، هناك انخفاض آخر في الإمكانات السلبية في الغشاء النووي (-13 mV).
تشير هذه النتائج إلى أن الغلاف النووي ربما يكون حاجزًا لانتشار الأيونات الصغيرة مثل K + أو Na + أو Cl. في المظروف النووي الموجود في الأجاجيات ، ومع ذلك ، لا توجد إمكانية محتملة ، مما يشير إلى التبادل الحر للأيونات بين النواة والسيتوبلازم.

الآلية التي يتحقق بها مرور هذه المواد غير معروفة. ومع ذلك ، كشفت الدراسات الكيميائية الخلوية عن وجود ATPase في المسام التي قد توفر الطاقة اللازمة لنقل الجزيئات الكبيرة.
أصل الغشاء النووي :
يعتبر الغشاء النووي بمثابة بنية سيتوبلازمية متخصصة ناشئة عن نظام الغشاء السيتوبلازمي. خلال الطور البعد ، يتم تكوينه عن طريق تراكم حويصلات الشبكة الإندوبلازمية حول المجموعة الصبغية ، والتي في وقت لاحق على الصمامات لتشكيل غشاء كامل.
هناك نظريتان أساسيتان حول كيفية إعادة تشكيل الغلاف النووي بعد الطور ، إما أن يقوم الصفيحة بتجميع النوفو في المواقع التي تتلامس فيها الكروموسومات والسيتوبلازم مع بعضها البعض (جونز 1960) أو أجزاء من الشبكة الشبكية الداخلية الإكلينيكية الموجودة بالفعل. نواة الطور البعدي ودمجها لتشكيل الغلاف النووي.
في الخلايا المنوية للجراد ، على سبيل المثال ، فإن حويصلات 0.5 which التي تشكلت على سطح الكروموسومات ، لا يمكن تمييزها عن حويصلات الشبكة الإندوبلازمية (بحر ، 1959 ، 61). يرتبون أنفسهم تدريجياً لكي يوازي سطح النواة ثم يندمجون لتشكيل غلاف نووي مستمر.
2. النسغ النووي:
إن كتلة "karyolymph" أو "النسغ النووي" ، وهي كتلة غير مضبوطة أو خفيفة من مادة acidophilic ، تملأ بالكامل الفضاء النووي الذي توجد فيه المكونات الأخرى. كان كلود هو الذي عجل في عام 1943 بالصفراء النووية بواسطة تثبيتي هيستولوجي وملطخ بالأصباغ الحمضية. في البيض والخلايا الكبيرة (على سبيل المثال ، Acetabularia) ، فإن النسغ النووي مرئي بشكل واضح. وفقا لالصقة (1951 ، 58) فإنه يعطي اختبار cytochemical إيجابية للجيش الملكي النيبالي ، بما في ذلك البروتينات SH ومجموعات البروتينات السكرية.
وقد ثبت أيضا العديد من المسارات الأيضية في النسغ النووي ، على غرار السيتوبلازم. وتشمل هذه التحلل السكري ، التحويلة الفوسفاتية hexose ، دورة حمض الستريك ، إلخ. تحويلة الفوسفات الأحادي السداسي مهم لأنه يوفر النواة مع البنتوز. علاوة على ذلك ، في النواة NAD + يتم توليفها والتي هي أنزيم من العديد من dehydrogenases. الأنزيمات المميزة للنواة هي الحمض النووي و polymerases الحمض النووي الريبي.
التركيب الكيميائي للنواة:
تم تحليل المكونات الكيميائية التالية من المواد النووية المعزولة:
(ط) البروتينات النووية
(أ) النيوكليو بروتامين
(ب) النوكليونات
(ج) غير الستيرون أو البروتينات الحمضية.
(2) الأحماض النووية.
(3) الانزيمات.
(د) الدهون.
(ط) Nuceoproteins:
وبصرف النظر عن الأحماض النووية ، فإن المكونات الأخرى المهمة لنواة الخلية هي مكونات البروتين. تتكون كروموسومات النباتات والخلايا الحيوانية بخلاف المادة الجينية للبكتيريا من الحمض النووي المرتبط بالبروتينات. في الكروموسومات المتعددة للكروموسومات العملاقة تم العثور على البروتين في مناطق النطاق و integrand من نشاط الجينات.
يرتبط البروتين مع الحلقات الممتدة من صبغيات اللمعان. يحتوي النيوكليلي على نسب عالية من البروتين ، وهناك بالطبع بروتينات موجودة في النسغ النووي أو النواة ، وبالاشتراك مع الدهون ، في الأغشية النووية.
جزء البروتين في النواة هو بالتأكيد معقد للغاية وله العديد من المكونات. من بين هذه الأنواع الأكثر شهرة هي بروتينات بسيطة للغاية وبسيطة: البروتامين والتاريخ. بالإضافة إلى هذه البروتينات الحمضية ، فإن ما يسمى بالبروتينات والأنسجة غير الستيرون موجودة أيضًا.
(أ) بروتامين أو نوكليو بروتامين:
كان Miescher الذي اكتشف بروتامين في الحيوانات المنوية في سمك السلمون. هذه هي بروتينات بسيطة بسيطة لها وزن جزيئي منخفض جدا. هذه هي غنية جدا في arginine الأحماض الأمينية الأساسية وتوجد في الحيوانات المنوية من بعض الأسماك وتحيط بإحكام على الحمض النووي عن طريق الربط الملح.
وبما أن البروتامين أصغر من الهستونات ، مما يعني أن الكروموسومات والنواة يمكن أن تكون معبأة في مساحة أصغر ، مما يسمح بقدر أكبر من الحركة للحيوانات المنوية. بشكل عام ، فهي تتكون من حوالي 28 عديد ببتيد أرجينين ، يبلغ طولها الإجمالي 100 درجة وتحتوي على 19 أرجينين و 8 أو 9 أحماض أمينية غير أساسية. أثناء التطوير ، هناك استبدال تدريجي للهستونات بواسطة بروتامين. قد يكون هذا نتيجة لارتفاع تقارب البروتامين في الحمض النووي.
(ب) مستعمرات النوى :
تم اكتشاف histones في كريات الدم الحمراء أوزة بواسطة Kossel وبعد فترة وجيزة تم العثور عليها في الغدة الصعترية من قبل Lilientfeld. كما تم العثور على histones في نواة خلية نباتية معينة ، وخاصة جنين القمح والأرز من قبل Mirsky.
والبروتينات النواة هي أيضًا بروتينات أساسية لها وزن جزيئي يبلغ حوالي 12000. بالإضافة إلى حوالي 13٪ أرجينين ، تحتوي الهستونات على بقايا أساسية أخرى بما في ذلك الليسين والحامض. تم عزل عدة هيستون من تركيبة مختلفة ، وتم وصف ثلاثة أنواع.
(ط) غنية جدا بالليسين.
(الثاني) الارجينين الغنية.
(ثالثا) وغني قليلا يسين.
هذه هي غير متجانسة وتتألف من عدة مكونات. توجد الهستونات في جميع نويات الكائنات الحية العليا ، على الرغم من أنها لم تدرس إلا القليل في النباتات. غير أن تلك الموجودة في النباتات تشبه الهستونات الموجودة في الفقاريات.
إن الوظيفة الرئيسية للـ هيستون المنسوبة هي وظيفة "الصمغ" الكروموسومي الذي يربط بين الوحدات الوراثية للحمض النووي. ومن المعروف أيضا أن الحمض النووي وحده ، وأن البروتينات (أساسا histones) في مجمع البروتينات النووية يمكن أن تحمي جزئيا الحمض النووي من التلف الإشعاعي.
تشير الأدلة الآن ، مع ذلك ، إلى أن الدور المهم الذي تلعبه الهستونات يكمن في تمثيل النشاط الوراثي للخلايا. اقترح ستيفن وستيدنيان في عام 1950 أن الهستونات تتفاعل مع الحمض النووي بطريقة معينة ، مما يمنعها من العمل كصفيحة لتخليق الحمض النووي الريبي ، وبالتالي منع منع نقل المعلومات الوراثية إلى السيتوبلازم.
(ج) غير هيستونيس أو بروتينات حمضية :
وأدى التحليل الكيميائي لخيوط nuceli و Chromatin المعزولة إلى إظهار نوع آخر من البروتين والذي يطلق عليه عموما البروتين غير الهستوني. يحتوي هذا البروتين على التربتوفان وله خصائص حمضية.
تم تحديد جزء آخر غير هيستوني من خيوط الكروماتين المعزولة. تظهر البقايا غير القابلة للذوبان تحت المجهر كخيوط حلزونية تحافظ على خصائص الكروموسومات الطورية. من وجهة نظر كيميائية ، من المثير للاهتمام أن هذه الكروموسومات المتبقية تحتوي على حمض ريبونوكلييك أكثر من الحمض النووي الريبي.
كمية كبيرة من البروتينات غير الستيرون الموجودة في الخلية النشطة الأيضية تتناقض بشكل ملحوظ مع تكوين الحيوانات المنوية ، والتي هي أقل نشاطا بكثير.
(2) الأحماض النووية:
الأحماض النووية ، والتي هي من نوعين: حمض الريبونوكلييك (RNA) وحامض الديوكسي ريبونوكلييك (DNA) وقد لوحظت في المنطقة النووية للخلية.
(3) الانزيمات النووية:
تم العثور على عدد من الانزيمات في الحويصلات الجرثومية معزولة من البويضات الضفدع من قبل Brachet و Dupsiva. من الصعب التأكد من تركيزها الفعلي بسبب النفاذية العالية للغشاء النووي ، كما أن الإنزيمات عرضة للتسرب من الحويصلات الجرثومية المعزولة أثناء عزلها.
وفقا لدراسات حديثة فإن الانزيمات النووية تنقسم إلى فئتين. البعض لديهم توزيع عام حيث يوجد البعض الآخر بشكل تفضيلي في بعض الأنسجة. في المجموعة الأولى ، تم العثور على تركيز عالي فقط لبعض الإنزيمات المتعلقة بعملية التمثيل الغذائي للنواة ، مثل الأدينوزين دياميناز ، فوسفوريلاز النوكليوسيد والجوانيز.
الانزيمات الأخرى مثل esterases موجودة في تركيز متفاوت. لا يزال آخرون ، مثل الفوسفاتيز القلوية ، الفوسفاتاز النوكليوتيد و β-glucoronidase إما موجودة في تركيز منخفض أو تفتقر تماما. من الانزيمات الخاصة ، يبدو أن الكاتلاز وأرجيناز تتركز في بعض النوى ولكنها تفتقر إلى غيرها.
(4) الدهون النووية:
وقد تم التحقيق في محتوى الدهون من النواة في نواة معزولة. في الآونة الأخيرة تم الإبلاغ عن أن مجمع بروتين ليبو في النوى من طحال الثيران وكريات الدم الحمراء ما يقرب من 10 ٪ من الدهون ، وهذه تعطي اختبارات إيجابية ل phospholipids والكولسترول.
3. النواة:
تحتوي النواة على حبيبة كبيرة وكريحية وحمضية كثيفة تعرف باسم النيكل. اكتشفت Fontona النيكليولي في عام 1781. وقد ثبت في القرن التاسع عشر أن حجم النواة مرتبط بالنشاط الاصطناعي للخلية.
لذلك ، فإن الخلية ذات النشاطات الاصطناعية الصغيرة أو منعدمة ، على سبيل المثال ، خلايا الحيوانات المنوية ، و blastomeres ، وخلايا العضلات ، وما إلى ذلك ، وجدت أنها تحتوي على نواة صغيرة أو بدون ، في حين تحتوي البويضات والخلايا العصبية والخلايا الإفرازية التي تجمع البروتينات أو المواد الأخرى بشكل نسبي. نواة كبيرة الحجم.
يعتمد عدد النواة في النواة على الأنواع وعدد الكروموسومات. قد يكون عدد النواة في النواة في الخلايا واحد أو اثنين أو أربعة. موضع النواة في النواة غريب الأطوار.
بنية دقيقة من nucleolus :
فيما يتعلق بالبنية الدقيقة للنواة ، أبلغ عنها كل من Estable و Sotelo (1955) أنهما يتكونان من جزأين - nucleolonema خيطية و pars amorpha. يقال أن النيوكليوليونما تقسم وتتوزع بالتساوي في انقسام فتات الابنة وتعتبر بنية دائمة تستمر طوال الانقسام الفتيلي بالترافق مع الكروموسوم.

ويعتقد أن الجزء الخيطي من النواة يحتوي على الحمض النووي في حين أن جزئيا amorpha تتكون جزئيا من الحمض النووي الريبي. يخضع Pars amorpha لدورة تكوين مميزة في الطور البيني و الاختفاء في الطور الأولي.
سمح إدخال وسائل وتقنيات تضمين جديدة بتحليل أفضل للتنظيم النووي.
وبالتالي يمكن التعرف على أربعة مكونات أساسية:
1. جزء fibrillar ما يقرب من 50A وقطرها تصل إلى 300-400 ألف درجة طويلة (Marrinozzi ، 1963). في بعض الحالات ، يمكن النظر إلى هذه الليفالات على أنها بنية مزدوجة تقطعت بهم السبل (Terzakis ، 1965). كما تم وصف هذه الهياكل كعناصر أنبوبي بقطر 15 A 0 .
2. جزء حبيبي مكوّن من حبيبات كثيفة بقطر يبلغ متوسطه 150-200 درجة مئوية ، أكثر أو أقل عدداً على طول وبين خيوط الشبكة الفايبرالية (Marrinozzi، 1963).
3. منطقة غير متبلورة ذات كثافة كهربائية منخفضة ، توجد في بعض النواة (Terzakis ، 1965) مصنوعة من البروتين.
4. إن الكروماتين المصاحب للنواة والموجود حول النواة وكثيراً ما يحتوي على مكونات داخل النواة لتكون ثابتة في خلايا الفقاريات ، ولكن قد تختلف مقاديرها أو كمياتها.
الكيمياء :
تشير الدراسات الكيميائية الخلوية إلى أن 5 إلى 10 بالمائة من النواة هي RNA. الباقي هو البروتين. مكونات البروتين الرئيسية هي الفوسفات. لم يتم العثور على نغماته في أنوية معزولة.
هناك أدلة تشير إلى وجود orthophosphate ، والتي قد تكون بمثابة مقدمة من الفوسفور RNA. لا يعرف إلا القليل عن محتويات الأنزيم في النواة. ومع ذلك ، فإن وجود الفوسفاتيز الحمضي ، و nucleoside phosphorylase و NAD + إنزيمات توليف قد أظهرت. وقد تم أيضًا ترجمة مثيلياز الحمض النووي الريبي في نواة خلايا معينة.
الحمض النووي غائب. قد تكون Nucleolus محاطة بحلقة من الكروماتين الموجب لـ Feulgen والذي يمثل في الواقع المناطق المتغايرة الصبغيات في الكروموسومات.
Biogenesis من Nucleolus أو دورة Nucleolar:
النواة كهيئة منظمة تفتقر إلى الاستمرارية. يختفي في بداية الانقسام الخلوي (الطور الأولي) ويظهر مرة أخرى في نهاية انقسام الخلية في مرحلة الطور.
تتشكل النواة في منطقة محددة بواسطة كروموسوم واحد أو أكثر من مجموعة الكروموسومات أحادية الصيغة الصبغية. تُعرف هذه الكروموسومات باسم الكروموسومات النووية.
غالبية الأنواع ثنائية الصبغيات تمتلك اثنين من الكروموسومات النووية في كل خلية ثنائية الصبغية أو الجسدية. ولكن في كروموسومات الإنسان التي يبلغ عددها 13 و 14 و 15 و 21 و 22 تشارك في تكوين النواة.
وتعرف المنطقة المحددة من هذه الكروموسومات النشطة في التكوين النووي باسم منطقة النيوكليولار النوكليوي. في كثير من الأحيان ولكن ليس دائمًا ، يتم تمييزه عن طريق تقييد ثانوي. ويحمل المنظم النووي الجينات اللازمة للـ RNA 18S و 28 S الريبوسيوم Ribosomal.
الانقباض الثانوي مع منظم nucleolar يختلف شكليا من تقلص ثانوي آخر. في مرحلة pachytene من Zea mays ، ترتبط هيئة تنظيم النوى الملطخة بالظلام مع المنطقة النووية.
أنواع الأنوية :
على أساس توزيع الحبيبات الريبوتينية والحبيبية يمكن تمييز ثلاثة أنواع من النيوكليلي (ويلسون).
1. Nuclcoli مع nucleolonemas ، التي توجد في معظم الخلايا تسمى plasmosomes.
2. نوكليولي مدمج بدون نيوكليولونيماس ، والتي تم وصفها في الغدة اللعابية من sciurid وفي protozoan ، Tetrahymena pyriformis. في هذه النيوكليولي يتم توزيع حبيبات ريبونوكليروتين والألياف بشكل منتظم.
3. نواة على شكل حلقة مع حبيبات ribouncleoprotein و ليفات موجودة فقط في المنطقة الطرفية (nodolonema الطرفية) تم الإبلاغ عنها في الخلايا البطانية ، خلايا العضلات الملساء وخلايا سرطان اللمفاوية. في هذه النواة ، تتكون المنطقة الوسطى من الكروماتين بشكل كبير.
المهام:
(ط) إنتاج الحمض النووي الريبي :
النواة هي واحدة من أكثر المواقع نشاطًا في تخليق الحمض النووي الريبي. وتنتج حوالي 70-90 في المئة من الحمض النووي الريبي الخلوي في العديد من الخلايا. وهو مصدر للجيش الملكي النيبالي الريبوزي (rRNA). يحتوي الكروماتين في النواة على جينات أو دنا ريبوسومي (rDNA) لتشفير الرنا الريبوسومي.
تمثل الليفرات أصل الرنا الريباسي والحبيبات في المرحلة التالية. الحبيبات في المقابل هي سلائف الريبوسومات. وهكذا ، فإن النيوكليول يصنع سلائف ريبوزومية بدلاً من ريبوسوم كامل.
كرومات ← Fibrils ← حبيبات ← ريبوسوم
(تحتوي على الحمض النووي) (التي تحتوي على الحمض النووي الريبي) (التي تحتوي على الحمض النووي الريبي).
قد تنتج النواة أيضًا بعض أنواع الرنا المرسال (mRNA) ، ونوعًا واحدًا على الأقل من الحمض النووي الريبي منخفض الجزيئ.
(2) تركيب البروتين :
اقترح ماجيو (1960) وآخرون أن تخليق البروتين يحدث في النواة. إذا كان هذا صحيحًا ، يتم إنتاج البروتينات الريبوسومية في النواة. دراسات أخرى ، ومع ذلك ، تشير إلى أن syntesized البروتينات الريبوسوم في السيتوبلازم.
(ثالثا) تشكيل ريبوسوم :
في مجموعة الكائنات حقيقية النواة يحتوي الترميز الجيني لـ RNA على سلسلة تحتوي على ما لا يقل عن 100 إلى 1000 نسخة مكررة من DNA. يتم إعطاء هذا الحمض النووي في شكل حلقات من الألياف الصبغية. ترتبط الحلقات الدنا مع البروتينات لتشكيل nucleoli.
الحمض النووي بمثابة قالب ل rRNA 45S. يتم تقسيم نصف الحمض الريبي النووي الريبي 45S إلى الأسفل لتكوين 28S و 18S rRNA. النصف الآخر مقسم إلى مستوى النوكليوتيد. داخل nucleolus يتحد الـ 28S rRNA مع البروتينات المصنوعة في السيتوبلازم لتشكيل الوحيدات الريبوسومية 60S. يربط الـ 18S rRNA أيضاً البروتينات لتشكيل الوحيدات الأربعين من الريبوسوم.
4. شبكة الكروماتين :
يظهر الكروماتين كهيكل شبيه بالهيكل ، ملفوف وممدود. هي ملطخة هذه الأصباغ الأساسية مثل وصمة Feulgen ، acetocarmine الخ ، وهذا هو السبب في هذه تسمى بألياف الكروماتين أو مادة الكروماتين (Gr. ، chromecolour ، soma = body).
هذه مرئية خلال مرحلة الطور البيني. خلال التقسيم الخلوي ، تصبح هياكل سميكة تشبه الشرائط المعروفة بالكروموسومات.
أهمية النواة:
تجربة هامرلينج:
أثبت J. Hammerling ، عالم الأحياء الألماني ، تجريبيا أهمية النواة. إذا تم قطع جزء يحتوي على النواة من Acetabularia من أحد الأنواع ، التي تتميز مورفولوجيا معينة ، فإن الشظية ستجدد خلية كاملة من هذا النوع.
هذه القدرة التجديدية تسمح بتجارب من النوع الموضحة في الشكل 8.6 حيث يتم الجمع بين نواة نوع واحد مع السيتوبلازم من أنواع مختلفة. أجرى هامبلرنج تجارب معينة باستخدام نوعين من الطحلب الأخضر ، Acetabularia. النوعان ، وهما A. crenulated و A ، البحر الأبيض المتوسط المستخدم في هذه التجربة يختلفان في شكل القبعات. بينما في A. crenulata سقف له أشعة فضفاضة ، في A. mediterranea يتم العثور على غطاء يشبه مظلة.

تقع النواة في كلا النوعين في جذع في أسفل الساق. إذا تم قطع الغطاء ، فسوف يتطور مرة أخرى وسيكون شكله من النوع الأصلي. ومع ذلك ، إذا تم إزالة التطعيم من نوع واحد بعد إزالة القبعات ، على الساق المجهرية (التي تحتوي على النواة) من الأنواع الأخرى ، سيتم تحديد شكل الغطاء عن طريق النواة وليس بالساق. إذا كانت النواة تنتمي إلى A. crenulata ، فإن شكل الغطاء سيكون من نوع crenulata. إذا كانت النواة تأتي من A. mediterranea ، فإن الغطاء يكون من نوع mediterranea.
عندما تكون النواة موجودة ، سيكون شكل الغطاء متوسطًا. الاستنتاج المستخلص من هذه التجارب هو أن النواة تنتج مادة تدخل السيتوبلازم وتشارك في التحكم في نمو الخلايا وتشكل الخلية.
النتيجة الحاسمة هي أن مورفولوجية الخلايا المتجددة تصبح عمليا مثل الشكل الذي يتم من خلاله أخذ النواة. في الشظايا الهجينة التي تحتوي على النواة من أحد الأنواع ومعظم السيتوبلازم من الأنواع الأخرى ، تستمر المادة السيتوبلازمية القديمة لفترة من الزمن وقد تؤثر على شكل الخلية. ولكن في نهاية المطاف ، ينضب هذا ويستبدل بمواد منتجة حديثًا من النواة.