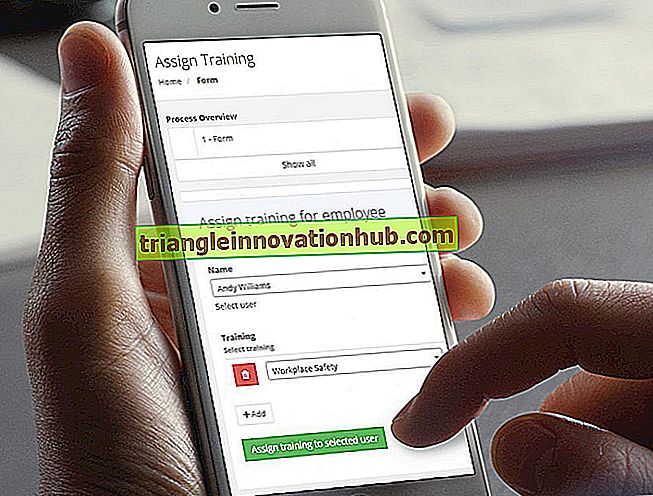
التركيب الجزيئي لغشاء البلازما
الهيكل الجزيئي لغشاء البلازما!
جميع الأغشية البيولوجية ، بما في ذلك غشاء البلازما والأغشية الداخلية لخلايا حقيقيات النواة ، لها بنية عامة مشتركة: فهي عبارة عن تجميعات لجزيئات البروتينات الدهنية والبروتينية التي تجمعها تفاعلات غير تساهمية.
يتم ترتيب جزيئات الدهون كطبقة مزدوجة مستمرة من 4 إلى 5 نانومتر سميك. يوفر هذا الدهن ثنائي الطبقة البنية الأساسية للغشاء ويعمل كحاجز كتيمة نسبياً لتدفق معظم الجزيئات القابلة للذوبان في الماء.
جزيئات البروتين "مذابة" في طبقة ثنائية الدهون وتتوسط الوظائف المختلفة للغشاء ؛ يخدم البعض لنقل جزيئات محددة إلى الخلية أو عرقها ؛ البعض الآخر عبارة عن إنزيمات تحفز التفاعلات المرتبطة بالغشاء ؛ ومازال البعض الآخر بمثابة روابط هيكلية بين الهيكل الخلوي للخلية والمصفوفة خارج الخلية ، أو كمستقبلات لاستقبال الإشارات الكيميائية ونقلها من بيئة الخلية.
جميع أغشية الخلايا هي هياكل ديناميكية وسوائل: معظم جزيئاتها الدهنية والبروتينية قادرة على التحرك بسرعة في مستوى الغشاء. الأغشية هي أيضًا هياكل غير متماثلة: تختلف تراكيب الدهون والبروتين في الوجهين عن بعضها البعض بطرق تعكس الوظائف المختلفة التي يتم إجراؤها على السطوح.
على الرغم من أن مكونات البروتين والدهون تختلف بشكل كبير من نوع واحد من الغشاء إلى آخر ، إلا أن معظم المفاهيم الهيكلية والوظيفية الأساسية تنطبق على الأغشية داخل الخلايا وكذلك الأغشية البلازمية.
بعد النظر في بنية وتنظيم المكونات الرئيسية للأغشية البيولوجية - الدهون والبروتينات والكربوهيدرات - سنناقش خلية الآليات التي تستخدم لنقل الجزيئات الصغيرة عبر أغشية البلازما وآليات مختلفة جدا التي تستخدمها لنقل الجزيئات الكبيرة والجسيمات الكبيرة عبر هذا الغشاء.
الدهون طبقة ثنائية:
أول إشارة إلى أن جزيئات الدهون في الأغشية البيولوجية منظمة في طبقة ثنائية جاءت من تجربة أجريت في عام 1925. تم استخراج الدهون من أغشية خلايا الدم الحمراء مع الأسيتون وطرح على سطح الماء. ثم انخفضت المساحة التي يشغلونها عن طريق حاجز متحرك حتى تم تشكيل فيلم أحادي الجزيئات (أحادي الطبقة).
احتلت هذه الطبقة الأحادية مساحة نهائية حوالي ضعف مساحة خلايا الدم الحمراء الأصلية ، لأن الغشاء الوحيد في خلية الدم الحمراء هو غشاء البلازما. وخلص المجربون إلى أن جزيئات الدهون في هذا الغشاء يجب أن يتم ترتيبها كطبقة ثنائية مستمرة.
كان الاستنتاج صحيحًا ، ولكن اتضح أنه يستند إلى افتراضين خاطئين كانا يعوضان عن بعضهما بعضا. من ناحية ، لم يستخلص الأسيتون الدهون الكاملة. من ناحية أخرى ، استندت المساحة السطحية المحسوبة لخلايا الدم الحمراء إلى المستحضرات المجففة وكانت أقل بكثير من القيمة الحقيقية في المستحضرات الرطبة.
لذلك ، كان للنتائج المستخلصة من هذه التجربة تأثير عميق على بيولوجيا الخلية ؛ ونتيجة لذلك ، أصبح طبقة الدهون الثنائية جزءًا مقبولًا في معظم نماذج بنية الغشاء ، قبل وقت طويل من تأسيسها.

دانييلي-دافسن أو بروتين-دهون بروتين أو ساندويتش
أشار هارفي وكول (1931) إلى وجود البروتين من خلال دراسة التوتر السطحي للخلايا. قاد هذا دانييللي ودافسون إلى اقتراح نموذج البروتين الدهني لغشاء الخلية. وفقا لهذا النموذج يتكون غشاء البلازما من طبقتين من جزيئات الدهون كما هو موضح في نموذج طبقة ثنائية الدهون.
تحتوي جزيئات الدهون على مناطقها القطبية على الجانب الخارجي. يعتقد أن بروتينات الجلوبيولين مرتبطة بالمجموعات القطبية للدهون. إن نهايات غير القطبية الكارهة للماء من طبقتين من الدهون تواجه بعضها البعض ، في حين ترتبط نهايات قطبية الماء مع جزيئات البروتين بالتفاعل الكهروستاتيكي. المسام القطبية المرتبطة بالبروتين موجودة في الغشاء. وتتكون هذه المسام من الاستمرارية الدورية للطبقات الخارجية والداخلية لبروتينات غشاء البلازما.
تعديلات على غشاء Danielli-Davson Model:
تم وصف العديد من التعديلات على الترتيب أعلاه:
(أ) قامت بعض أغشية البلازما بطباعة سلاسل ß من البروتينات على سطوح طبقة الدهون الثنائية.
(ب) α سلاسل ملفوفة من البروتين الحلزوني على سطح طبقة ثنائية الدهون.
(C) مع البروتينات الكروية على كل من السطوح.
(د) مع بروتينات مطوية على كل من السطوح والبروتينات حلزونية تمتد إلى المسام.
(E) مع بروتين ß-cham مطوي على جانب واحد والبروتين كروي على الجانب الآخر.


نموذج غشاء وحدة روبرتسون:
تم طرح نموذج غشاء الوحدة في عام 1953 أثناء دراسة الخلية تحت المجهر الإلكتروني. تم اعتبار هيكل الغشاء الأساسي للوحدة عامًا لمجموعة متنوعة واسعة من الخلايا النباتية والحيوانية. جميع عضيات الخلية مثل جسم Golgi ، والميتوكوندريا ، والشبكة الإندوبلازمية ، والغشاء النووي وما إلى ذلك ، لها بنية غشاء الوحدة.
يعتبر غشاء الوحدة ثلاثي الطبقات مع طبقة شحمية ثنائية الجزيئات بين طبقتين من البروتين. طبقتان متوازيتان خارجيتان من الطبقة العميقة من 20A ° تتوافقان مع طبقتين من البروتين. وتبلغ الطبقة المتوسطة المتوسطة السميكة حوالي 35 درجة مئوية في سمك مماثل لسلاسل الهيدروكربون من الدهون.
وبالتالي فإن غشاء الوحدة هو حوالي 7A درجة في السمك. في هذا الصدد ، يعيد تجميع نموذج دانييللي-دافسن. ومع ذلك ، فإنه يختلف عن نموذج دانيزلي-دافسون في أن البروتين غير متماثل. على السطح الخارجي هو mucoprotein ، بينما على السطح الداخلي هو بروتين غير mucoid.

اعتراضات على غشاء الوحدة
ازدادت الاعتراضات على نموذج غشاء الوحدة خلال الستينات ، مما أدى إلى إعادة فحص تفاعلات البروتين الدهني وإلى نماذج جديدة شددت الدراسات التي أجريت على FS Sjostrand (1963) على الأغشية الشبكية الإندوبلازمية والميتوكوندريا والكلوروبلاست على الفروق بين الملامح المرصودة للأغشية. والتوحيد الذي كان مطلوبًا بمفهوم الوحدة الغشائية.

تحتوي أغشية الميتوكوندريا والكلوروبلاست على وحدات من الجسيمات في أو على الغشاء. لم يقدم غشاء البلازما نفس مظهر أغشية الميتوكوندريا أو البلاستيدات الخضراء. يبدو أن هناك حاجة إلى نماذج مختلفة لوصف أنواع وظيفية مختلفة. أصبح هذا النهج غير المناسب غير ضروري عندما تم اقتراح نموذج غشاء الفسيفساء.
نموذج غشاء أكبر:
مثل النموذج الثلاثي هنا أيضا طبقة الدهون تقع بين طبقتين من البروتينات الهيكلية. وصف روبرتسون الطبيعة المختلفة للأسطح الخارجية والداخلية للغشاء. كان يعتقد أن السطح الداخلي مغطى بالبروتين غير المقترن والسطح الخارجي مع بروتين سكري ، والذي يتم تركيبه على سلاسل البروتين oligosaccharide الهيكلي مع محطات حمض السياليك المشحونة سلبيا تعلق على بروتين سكري.
نموذج Micellar:
وقد تم تفسير تفسير بديل للبنية الجزيئية لغشاء البلازما من قبل هيلير وهوفمان (1953). وقد اقترحوا أن الأغشية البيولوجية قد يكون لها نمط غير صفائحي ، يتكون بدلا من فسيفساء من وحدات فرعية كروية تعرف بميسيليس ، والتي تحتوي على قلب للدهون وقشرة محبة للماء للمجموعات القطبية.
من المحتمل أن تكون micelles lipid كتل بناء للأغشية لأنها تميل نحو الارتباط العفوي. في هذا النموذج من التركيب الغشائي يمكن أن تشكل مكونات البروتين في الغشاء أحادي الطبقة على جانبي مستوي الميكروفونات الدهنية.
يمكن استبدال وحدات فردية من الفسيفساء micellar بجزيئات الإنزيمات الفردية أو عن طريق صفائف من الانزيمات مع منظمة ثلاثية الأبعاد دقيقة ، مما يتيح وظائف محددة لتكون "بنيت في" إلى بنية الغشاء.
ويعتقد أن الفراغات بين المذيلات الكروية تشكل مسام مملوء بالماء 0.4 نانومتر (4A °) في القطر ، مبطنة جزئياً بالمجموعات القطبية للمذبلات وجزئياً بالمجموعات القطبية لجزيئات البروتين المرتبط.

نموذج فسيفساء السوائل:
اقترح هذا النموذج سنجر ونيكلسون (1972). وفقا لهذا المفهوم ، يتم ترتيب جزيئات الدهون لتشكيل طبقة ثنائية مستمرة التي تشكل العمل الإطار الهيكلي لغشاء البلازما. يتم ترتيب جزيئات البروتين بطريقتين مختلفتين. توجد بعض البروتينات بشكل حصري متاخمة للأسطح الخارجية والداخلية للطبقة الثنائية الدهنية وتسمى بروتينات خارجية. بروتينات أخرى تخترق طبقة ثنائية الدهن جزئيا أو كليا وتشكل بروتينات مدمجة أو داخلية.

الدهون والبروتينات المتكاملة لغشاء البلازما هي أمفيتيكية في الطبيعة. مصطلح "أمفيباتي" صاغه هارتلي (1936) لتلك الجزيئات التي تحتوي على مجموعات مسعورة ومحببة للماء. تميل الجزيئات الأمفيمية إلى أن تشكل مجاميع بلورية سائلة توجد فيها مجموعات مسعورة أو غير قطبية داخل الطبقة الثنائية ، وتوجه مجموعات محبة للماء نحو مرحلة المياه. لذلك ، تشكل جزيئات الدهون طبقة ثنائية مستمرة.
يتم تقطيع البروتينات المتكاملة في طبقة ثنائية الدهون ، مع مناطقها القطبية البارزة من السطح والمناطق غير القطبية المضمنة في طبقة ثنائية الدهن. يشرح هذا الترتيب لماذا تتعرض المواقع النشطة للأنزيمات والبروتينات السكرية المستضدية للسطح الخارجي للأغشية. يوضح هيكل quasifluid لغشاء البلازما حركة كتلة جزيئات البروتين ذات الحجم الكبير عبر الغشاء.
المسام في غشاء البلازما:
غشاء البلازما مثقب من قبل المسام. يبلغ قطرها حوالي 0. 35 نانومتر (نانومتر) ، أكبر قليلاً من أيونات الصوديوم. أقل من 0.1 في المائة من غشاء البلازما مثقب من قبل المسام ، في حين أن 99.9 في المائة من سطح الخلية غير منفذ للأيونات. وقد اقترحت عدة نماذج لهياكل المسام.
البعض منهم:
1. المسام الهيكلية :
هذه هي الثقوب الأسطوانية الدائمة التي تقاوم ورقة طبقة ثنائية مستمرة.
2. المسام الديناميكية:
هذه المسام هي ثقوب أسطوانية مؤقتة بدلاً من كونها دائمة. تظهر فقط في وقت المدخول.
3. رصف مسام القناة :
ووفقًا لهذا المفهوم ، يُنظر إلى المسام على أنها زوايا كتل الرصف السداسية المملوءة تقريبًا والمكونة من الوحدات الفرعية للدهون والبروتينات.
4. مسام قناة البروتين :
تعتبر هذه المسام أجزاء من نموذج الفسيفساء بروتين الدهون. تشكل هذه القنوات الصغيرة لبروتينات معينة مدمجة في الغشاء الذي يمكن من خلاله أن تنتشر الأيونات والجزيئات الصغيرة.
5. ايونوفور:
الأيونوفور هي عبارة عن polypeptides صغيرة ذات نهاياتها مسعور ماء وغيرها. يذوب الطرف الغضروف (الخارجي) في الغشاء بينما يلتقط الطرف المائي (الجانب الداخلي) المواد الأيونية أو المواد القابلة للذوبان في الماء ويقذفها على الجانب الآخر. تساعد الأيونوفورز في تبادل المواد من أو داخل الخلية.
التخصص أو التعديلات :
مع زيادة دقة المجهر الإلكتروني تم التعرف على التخصص العديد من سطح الخلية (Sjostrand 1956 ؛ Fawcett 1958). يلي وصف Fawcett (1958) يتناول التخصصات المختلفة من غشاء البلازما درس طبوغرافيا.
زغيبات:
في الظهارة microvilli المعوية بارزة جدا وتشكل بنية مدمجة التي تظهر تحت المجهر الضوئي كحدود مخططة. وتمثل هذه الميكروفيلي ، التي يتراوح طولها بين 0.6 و 0.8 ميكرومتر وقطرها 0.1 ميكرومتر ، العمليات السيتوبلازمية التي يغطيها غشاء البلازما. داخل النواة السيتوبلازم يتم ملاحظة الميكروفيلمونات الدقيقة التي تشكل في السيتوبلازم شبكة طرفية.
يتم تغطية السطح الخارجي لل microvilli بواسطة طبقة من المواد الخيطية (معطف غامض) تتألف من جزيئات ميكروية بروتين جليكو بروتين. Microvilli زيادة سطح امتصاص فعالة. على سبيل المثال ، قد تحتوي الخلية الواحدة على 300 ميكروفيلي وفي ميلليمتر مربع من الأمعاء قد يكون هناك 200 ، 0 ، و 000. تشكل المسافات الضيقة بين الزغيبات الصغيرة نوعًا من الغربال الذي يجب أن تمر خلاله المواد أثناء الامتصاص.
العديد من الخلايا الأخرى ، بالإضافة إلى ظهارة الأمعاء ، لديها زغابات صغيرة ، على الرغم من عدد أقل. وقد تم العثور عليها في الخلايا الظهارية ، في الخلايا الظهارية من غشاء المثانة ، الرحم والكيس المحي ، في الخلايا الكبدية وهكذا دواليك.
حدود الفرشاة في أنبوب الكلى مشابهة للحدود المخططة ، على الرغم من أنها ذات أبعاد أكبر. مادة غير متبلورة بين microvilli يعطي رد فعل شيف الحمضي الدوري ل polysaccharides. بين microvilli ، في القاعدة ، وغشاء الخلية يخترق في السيتوبلازم قمي. هذه الابتزازات هي على ما يبدو المسارات التي تدخل كميات كبيرة من تدفقها من خلال عملية مشابهة لالإبروكسيات.

Desmosomes أو Macula Adherens:
Desmosomes هي تقاطعات الخلايا الموجودة بشكل أساسي في خلايا ظهارة عمودي بسيطة .. تحدث هذه كمناطق متخصصة على طول السطوح التلامسية. تحت المجهر الضوئي ، ينظر إلى الديسموسومات على أنها أجساد ملطخة بدقّة. تحت المجهر الإلكتروني تظهر هذه على شكل ثخانة تشبه الزر على السطح الداخلي لأغشية البلازما للخلايا المجاورة عند نقطة الاتصال.
يتم اجتياز الأطياف من خلال الألياف الليفية السيتوبلازمية الدقيقة التي تدعى الليف الليفي ، والتي تشكل نوعًا من الحلقة في القوس العريض. هذه الخيوط تثبت التقارب وتعمل كمواقع تثبيت للبنى السيتوبلازمية. يتم فصل أغشية البلازما من الخلايا المجاورة في مناطق desmosomes بواسطة الفضاء بين الخلايا حوالي 30-35 نانومتر. وهي مليئة بمواد طلاء كثيفة متداخلة تشكل خطًا داكنًا في المنتصف. وهي مكونة من عديدات السكاريد المخاطية والبروتينات.
تهتم ال desmosomes في المقام الأول بالتصاق الخلية ، ولكنها تساعد أيضًا في الحفاظ على شكل الخلية ، مما يوفر لها الصلابة والدعم الخلوي. يتم الحصول على الأول عن طريق مادة الطلاء بين الخلايا والأخيرة من اللودات.

Plasmodesmata :
في بعض الأحيان ، يتم ربط الخلايا بجسور السيتوبلازم التي تمر بين مسام جدار الخلية أو غشاء البلازما بين الخلايا المجاورة ، وتسمى هذه الصلات بـ plasmodesmata. عادة ما تكون بسيطة ولكنها قد تجد أيضا plasmodesmata astomosing. توزيعها ورقمها قد يكون أيضًا كبيرًا جدًا. تم اكتشافهم من قبل تانغ! (1879) وتم تسميته على هذا النحو من قبل Strasburger (1882).
غالبًا ما ترتبط الشبكة الإندوبلازمية ارتباطًا وثيقًا بسطح الخلية عند النقاط التي توجد فيها البلاسميات. من خلالها يتم الاحتفاظ باستمرار الاستمرارية السيتوبلازمية بين الخلايا المجاورة. أنها توفر وسيلة للتفاعل بين الخلايا المتجاورة التي يتم فصلها في مناطق أخرى. من خلالها يمكن أن تنتقل المواد من خلية إلى أخرى.
من غير المعروف ما إذا كانت جميع البلاسموديماتا مشابهة لبعضها البعض. هناك بعض الاختلاف لأنها لا تنتج فقط في الوقت الذي تنقسم فيه الخلايا بل تتشكل أيضًا بشكل عفوي بين الخلايا التي نمت في اتصال مع بعضها البعض ، على سبيل المثال tiloses في عناصر وعاء الخشب.
قد تحدث منفردة أو يمكن تجميعها في مجموعات. في كثير من الجدران الأولية ، عادة ما ترتبط البلاسموديماتا بتخفيض مخفض لمواد الجدران ، وتعرف المنطقة بعد ذلك بالحفرة الأولية أو الحقل.

نصفي desmosomes:
توجد هذه في السطح القاعدي لبعض الخلايا الظهارية. هيكلها مشابه للديسموسومات ، لكن يتم تمثيلها بمقدار النصف. نظيرتها عادة ما يتم تمثيلها من قبل الألياف كوليجين.
أشرطة الطرفية :
تعرف قضبان المحطة الطرفية أيضًا باسم الوصلات الوسيطة أو الوصلات المنحدرة. تتشابه قضبان المحطات مع desmonomes إلا أنها تفتقر إلى اللدائن. في شريط المحطة الطرفية ، يكون غشاء البلازما سميكًا ويكون السيتوبلازم في المنطقة السميكة كثيفًا. تحدث القضبان الطرفية في الجزء الوسيط من غشاء البلازما للخلايا العمودية أسفل السطح مباشرة. الهوية الصحيحة ل zonula adhaerens لا تزال موضع شك (JP Trinkaus ، 1969).
تفاعلات الأغشية:
جانب آخر من أغشية الخلايا التي تستحق المناقشة هو التفاعل بين أغشية الخلايا المختلفة. التواصل بين الخلايا مهم في العديد من وظائف الخلية وخاصة أثناء تطوير الكائن الحي ، عندما تتفاعل الخلايا باستمرار مع الخلايا الأخرى.
قد تختلف طبيعة التفاعلات الغشائية من الجسور السيتوبلازمية الكاملة بين الخلايا إلى المناطق الموضعية في الوصلات الغشائية التي قد تشتمل على منطقة ملامسة صغيرة مثل بضعة أنجستروم أو كبيرة مثل عدة ميكرومترات. إن الطبيعة الهيكلية لجهة الاتصال الفعلية تقع عمومًا في واحدة من ثلاث فئات ؛ تقاطعات الفجوة ، الوصلات الضيقة ، وتقاطعات التفسخ.

تظهر الوصلات الفجوة كهيكل متعدد الطبقات عند ملاحظتها باستخدام المجهر الإلكتروني. ويبدو أنها أغشية لوحدتين متقاربة إلى بعضها البعض مع وجود فجوة بين 20 و 40 درجة مئوية. السماكة الكلية لتقاطع الفجوة بأكمله هي 170 إلى 190 درجة مئوية ، وهي موجودة في كل من الفقاريات واللافقاريات. لم يتم العثور عليها في ألياف العضلات والهيكل العظمي أو خلايا الدم الحمراء.

توجد تقاطعات ضيقة فقط في الفقاريات وتحدث في خلايا مثل الخلايا الظهارية. يبدو أن هذه التقاطعات عبارة عن اندماج حقيقي بين الغشاء ، وهي سميكة من 100 إلى 140 درجة مئوية.
تم العثور على تقاطب الحاشية فقط في اللافقاريات. وهي أكبر بكثير من الأنواع الأخرى من التقاطعات وتتميز بجسور متقاطعة إلكترونية كثيفة تمتد بين أغشية الخلويتين.