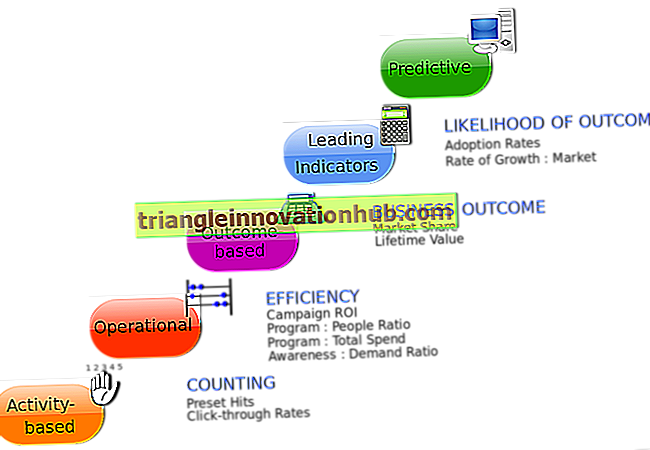
مراحل آلية تخليق البروتينات على ريبوسوم 70S
آلية تصنيع البروتين على 70S الريبوسوم تشمل المراحل التالية:
1. النسخ:
تبدأ عملية تخليق البروتين من قبل uncoiling من فروع جزيء الحمض النووي. أحد فروع جزيء الحمض النووي يعمل كقالب لتشكيل mRNA. يتم تشكيل الحمض الريبوزي النووي (mRNA) وفقًا للرموز الثلاثية للحمض النووي DNA عن طريق عملية النسخ أو النسخ.

الصورة مجاملة: images.fineartamerica.com/false-color-tem-of-ribosomes-photo-researchers-inc.jpg
حالما يتم تكوين الحمض النووي الريبوزي فإنه يترك النواة وتصل في السيتوبلازم حيث تعلق مع الوحيدات 30S من ريبوسوم. يحمل mRNA الكودونات الثلاثية لتخليق البروتينات.
2. مرفق مرنا مع 30S الريبوسوم وتشكيل polyribosome:
في الخلايا بدائية النواة لوحظ أنه قبل عملية تخليق البروتين تحدث الريبوسومات في حالة منفصلة وغير نشطة. يرتبط mRNA بالوحدة الريبوسومية 30S في وجود عامل بروتين F2. سرعان ما يأتي N-formylmethionyl-tRNA (F-met-t RNAf) من السيتوبلازم ويربط مع الشفرة الثلاثية الأولى (AUG أو AUA) من mRNA لبدء عملية تخليق البروتين وتشكيل مجمع البدء.
ويساعد تشكيل معقد البدء في GTP (Guanosine triphosphate) و 3 عوامل بروتينية (F 1 ، F 2 and F 3 ). بعد تشكيل معقد البدء ، توحد الوحدة الفرعية الريبوسومية 30S مع وحدة فرعية ريبوزمية 50S لتشكل الريبوسوم 70S. يحدث اتحاد الوحدات الفرعية الريبوسومية في وجود أيونات Mg ++ وعوامل F 1 و F 2 .
لا تتم قراءة رموز mRNA إلا بواسطة ريبوسوم واحد ولكن العديد من ريبوسوم يتحرك ويقرأ رموز mRNA. عندما يرتبط العديد من ريبوسوم مع مرنا يحدث تشكيل polysome أو polyribosome.


3. نقل الأحماض الأمينية إلى موقع تخليق البروتين:
يتم نقل الأحماض الأمينية من تجمع الحمض الأميني داخل الخلوي إلى ريبوسوم من قبل الحمض الريبي النووي النقال. تحدث عمليات النقل في مراحل مختلفة وهي كما يلي.
(أ) تفعيل الأحماض الأمينية:
كل من الأحماض الأمينية العشرين يحدث في السيتوبلازم في حالة غير نشطة. يتم تنشيط كل حمض أميني قبل ارتباطه مع الحمض الريبي النووي النقال الخاص به عن طريق إنزيم تنشيط معين يعرف باسم synthetase aminoacyl و ATP. تتفاعل الأحماض الأمينية الحرة مع ATP ، مما ينتج عنه إنتاج aminoacyl adenylate و pyrophosphate:
AA + ATP + enzyme → AA ~ AMP - enzyme + PP
الأحماض الأمينية aminoacyl synthetase aminoacyl adenylate Pyrophosphate synthetase complex
يرتبط منتوج التفاعل aminoacyl adenylate بالانزيم في شكل مركب أحادي التكافؤ. هذا estenylate amenacate adenylate المعقدة ثم esterifies لجزيئات الحمض النووي الريبي محددة. تحتوي الخلية على ما لا يقل عن 20 من إنزيمات الأمينازيل الانزيمية للأحماض الأمينية العشرين. كل إنزيم محدد ويرتبط بالحمض الأميني المحدد دون أي خطأ.
(ب) ملحق من الأحماض الأمينية المنشط إلى الحمض الريبي النووي النقال:
يظل الأدينيل أدينولايت مقيدًا بالانزيم حتى يصطدم بجزيء الحمض الريبي النووي النقال المحدد. بعد ذلك ، يتم نقل مجموعة الكربوكسيل لحمض بقايا الأحماض الأمينية للأمينوزيل أدينولات إلى مجموعة 3 OH من ريبوز الأدينوزين في نهاية CCA من الحمض الريبي النووي النقال. ونتيجة لذلك يتم تحرير AMP والإنزيم ويتم تكوين الحمض النووي الريبي النووي الأميني المنتج النهائي بواسطة الطريقة التالية:
'A' = موقع aminoacyl أو موقع فك التشفير ، 'P' = موقع peptidyl أو موقع تكثيف ، 'E' = موقع موجود ،
AA 1 = N-formyl methionine.
أأ 2 = ثريونين.
AA - AMP - Enzyme + tRNA → AA - tRNA + AMP + Enzyme
Aminoacyl adenylate and enzyme Aminoacyl — tRNA
يتحرك الأمينوئيل-ترنا إلى موقع تخليق البروتين ، أي ريبوسوم مرنا.
4. بدء تصنيع البروتين:
كما سبق أن ذكرنا أن بدء تخليق البروتين في بكتيريا الإشريكية القولونية (E. coli) ينطوي على تكوين مركب 70S. في ذلك ، يحتوي mRNA دائمًا على الكودون الثلاثي الأول كـ AUG في بدايته (أي نهاية 5)). الكودونات AUG هي أكواد لميثيونين الأحماض الأمينية.
الميثيونين لا يزال صياغيا وله دور مهم جدا في بدء عملية تصنيع البروتين. في كل نوع من أنواع البروتين يحتل الفورمول ميثيلون المرتبة الأولى في الجزيء وعندما يتم تصنيع جزيء البروتين تمامًا ، ينفصل ميثيل الميثيل في النهاية من جزيء البروتين المركب حديثًا عن طريق نشاط إنزيم hydrolytic.
منذ توليف البروتين ، تنمو سلسلة الببتيد دائمًا في التسلسل من مجموعة الأمينو الطرفية الحرة (—NH 2 ) نحو نهاية الكربوكسيل (—COOH) ، وظيفة formyl methionine -RRNA (AA 1 -RRNA في الشكلين 38.22 و 38.23). ) هو ضمان أن يتم توليف البروتينات في هذا الاتجاه.
في forme methionine - tRNA ، يتم حظر مجموعة amino (-NH 2 ) من قبل مجموعة formyl وترك فقط - مجموعة COOH المتاحة للتفاعل مع - مجموعة NH 2 من الحمض الأميني الثاني (AA 2 ). وبهذه الطريقة يتبع توليف سلسلة البروتين في التسلسل الصحيح.
5. استطالة سلسلة بولي ببتيد:
مع تكوين 70S الريبوسوم الوظيفي (أي ، 70S ، mRNA - F met tRNA) ، يتم جلب استطالة سلسلة polypeptide عن طريق الإضافة المنتظمة للأحماض الأمينية والحركات النسبية للريبوسوم و mRNA في وجود جزيئات GTP ، بحيث لا يزال هناك كودون ثلاثي جديد متوفر من أجل الحمض الريبي النووي النقال الجديد aminoacyl عند فك أو "A" موقع الريبوسوم في كل خطوة. وهكذا ، يجب أن يتحرك الحمض النووي الريبي المتناثر (F-met tRNA) من موقع فك الشفرة (موقع "A") إلى موقع peptidyl أو موقع "P" قبل أن يتراكم الحمض النووي الثاني من الحمض النووي الريبوزي (أي ، AA 2 tRNA) مع الكودون الثلاثي القادم الذي يحدث عند فك التشفير أو "A" موقع الريبوسوم.
يرتبط aminoacyl-tRNA (AA 2 tRNA) بكودون الموقع "A" بحضور GTP وبروتينين ، يطلق عليهما عوامل التحويل (المعينة Tn و Ts) والتي تظل مرتبطة بالريبوسوم. خلال عملية الربط هذه ، يتم تكوين معقد بين GTP ، وعوامل التحويل و aminoacyl tRNA (viz.، AA 2 -tRNA) ، والتي ترسب في نهاية المطاف aminoacyl tRNA في موقع A 'للريبوسوم مع إطلاق عوامل النقل-الناتج المحلي الإجمالي الفوسفات المعقد وغير العضوي.
في الخطوة التالية ، بسبب الحركة النسبية للريبوسوم و mRNA في وجود جزيء GTP واحد ، يصبح الكودون الثلاثي القادم (أي كودون UUU في التين .38.21 و 38.22) متاحًا لـ aminoacyl tRNA التالي (viz. AA 3 -tRNA) ) في موقع "A" للريبوسوم. في هذه المرحلة ، يحدث f met-tRNA عند مخرج موقع 'E' ، بينما يحدث AA 2 -tRNA في موقع peptidyl 'P'. الآن ، إنزيم يعرف باسم ترانسفيراز أنا أقوم بإطلاق الحمض الريبي النووي النقال من فورميل ميثيونين (f-met أو AA 1 ) وقلب الـ formyl-methionine إلى aminoacyl-tRNA (AA 2 -tRNA) المرتبط في موقع peptidyl أو 'P'. من المفترض أن يطلق العامل "G" الحمض الريبي النووي النقال الذي تم تصريفه أو نزعته من موقع "E" الخاص بالريبوسوم.
وهو يتبع المرحلة التالية من عملية الاستطالة والتي تشتمل على توليفة رابطة الببتيد عن طريق تفاعل بين المجموعة الأمينية الحرة للحمض الأميني القادم (أي ، AA 2 ) ومجموعة الكربوكسيل للحمض الأميني الأول (AA 1 ) وهو esterified إلى الحمض الريبي النووي النقال. يُطلق على الإنزيم الذي يحفز هذا التفاعل اسم الببتيديل ترانسفيراز (أو الببتيد سينتيثاز) وهو جزء لا يتجزأ من الوحدة الفرعية 50S. تستمد الطاقة لتوليف السندات الببتيد من انشقاق وصلة استر بين حمض أميني وحمض الريبوزي النووي.
وهكذا ، خلال استطالة سلسلة عديد الببتيد ، يدخل كل الحمض النووي الريبي المشحون أو الحمض النووي (Aminoacyl tRNA) في فك الترميز أو الموقع "A" ، ينتقل إلى موقع التكثيف أو "P" ، وينقل الحمض الأميني إلى نهاية الكربوكسيل من عديد الببتيد ، ينتقل إلى موقع الخروج ، حيث يتم نقل سلسلة polypeptide إلى الحمض الريبي النووي النقال المجاور على موقع التكثيف ، ثم يتم تحرير الحمض الريبي النووي النقال من الريبوسوم.
تعرف هذه العملية باسم translocation. ينتج عن هذا كودون ثالث قادم إلى موقع A ، وربط الحمض النووي الريبي (tRNA) الملائم بحمض أميني ثالث يرتبط بالموقع A. سوف تتكرر عملية تكوين السندات الببتيد والنقل.
وهكذا ، عندما يتحرك mRNA فيما يتعلق بالريبوسوم ، فإن جميع الكودونات سوف تتعرض في موقع ما وتنمو سلسلة الببتيد. يجب أن يحدث هذا التسلسل من الأحداث المتضمنة في الاستطالة بسرعة كبيرة لأنه قد تم حساب أنه ، E. coli تنمو في الظروف المثلى ، يمكن إنتاج سلسلة polypeptide من حوالي 40 حمض أميني في 20 ثانية.
6. إنهاء وإطلاق سلسلة بولي ببتيد:
عندما يتم الانتهاء من توليف سلسلة بولي ببتيد وفقا لكودونات مرنا ، ثم تتم عملية إنهاء وإطلاق سلسلة بولي ببتيد. يتم التحكم في إنهاء سلسلة polypeptide وإطلاق السلسلة المكتملة من الريبوسوم بعاملين.
يشار إلى إنهاء السلسلة من خلال ثلاثة توزيعات خاصة تنتهي في mRNA ، والتي في البكتيريا ، هي UAG ، UAA و UGA. هذه هي ما يسمى بالكودونات غير الحسّية التي لا ترمز لأي حمض أميني. ومع ذلك ، لا تزال سلسلة البولي ببتيد مرتبطة بحمض الري الحمض الريبي النووي الذي يرتبط بدوره بالحمض الريبوزي المرسال.
يتم إطلاق السلسلة من الريبوسوم تحت اتجاه ثلاثة بروتينات متميزة والتي تسمى عوامل الإطلاق وتسمى R1 R 2 و S. وتلتزم هذه الريبوسوم وتتحكم في التحلل المائي لربط الإستر بين الحمض الريبي النووي النقال وسلسلة البولي ببتيد. حالما يتم إنهاء السلسلة وإطلاقها ، يفصل الريبوسوم عن mRNA وينفصل إلى وحدتيه الفرعيين بسبب عمل عامل F 3 . إنه على استعداد لدخول دورة جديدة من توليف ببتيد.
7. تعديل polypeptide صدر:
تحتوي سلسلة polypeptide التي تم إصدارها على الميثيونين المصنوع في نهايته. يزيل إنزيم ديفورميلاز مجموعة الفورمين من ميثيونين. قد يزيل إنزيم إبيزبتيداز بعض الأحماض الأمينية من نهاية طرفية N أو نهاية الطرف C من سلسلة polypeptide.
في هذه المرحلة ، يمتلك البولي ببتيد (البروتين) هيكله الأساسي. على الأقل جزء من العديد من البروتينات له بنية ثانوية في شكل حلزون ألفا. قد تطوى السلسلة البروتينية مرة أخرى بنفسها ، وتشكل روابط داخلية (بما في ذلك تخزين روابط ثنائي كبريتيد) التي تثبت هيكلها الثلاثي في نمط مطوي بدقة وبشكل معقد في كثير من الأحيان. قد يتحد اثنان أو أكثر من الهياكل الثلاثية في بنية quarternary وظيفية. على سبيل المثال ، يتألف الهيموغلوبين من أربعة سلاسل متعددة البيبتيد ، واثنان متشابهان ، وأثنان من P -chains متطابقان. لا يصبح البروتين إنزيمًا نشطًا حتى يفترض نمطه الثالث أو الرباعي.
تنطوي عملية البدء والاستطالة والإنهاء على وساطة العديد من عوامل البروتين والتحلل المائي لـ GTP (غوانوسين ثلاثي الفوسفات) لتوفير الطاقة. تنطبق الخطوات التفصيلية التي تمت مناقشتها على المواد المخدرة ، مع بعض التعديلات أيضًا على الكائنات النواة.