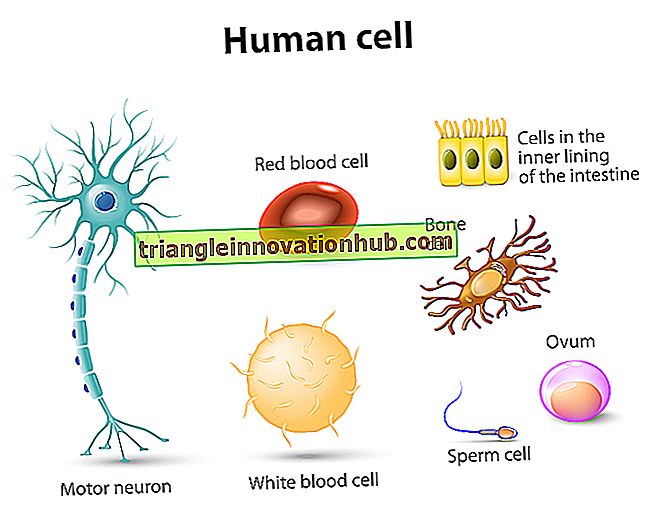
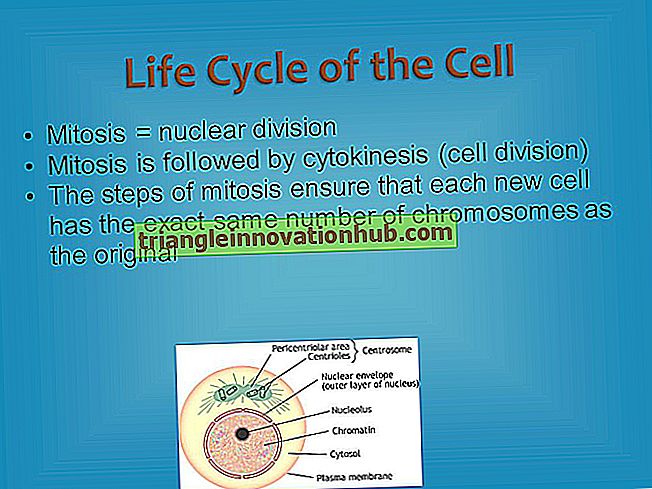
الريبوسومات: الحدوث ، التوزيع ، التركيب ، الأنواع ، التركيب الكيميائي والوظائف
الريبوسومات: الحدوث ، التوزيع ، التركيب ، الأنواع ، التركيب الكيميائي والوظائف!
الريبوسومات هي حبيبات basophilic موجودة في سيتوبلازم الخلية. بما أن هذه المادة لديها تقارب للبقع الأساسية الشبيهة بحبيبات الكروماتين في النواة ، فقد كان ذلك لفترة من الزمن تدعى مادة الكروميدية أو مادة الكروموفيل.
تمت ملاحظة الريبوسومات لأول مرة في الخلايا النباتية بواسطة روبنسون وبراون في عام 1953 أثناء دراسة جذور الفاصوليا مع المجهر الإلكتروني وبعد ذلك بفترة قصيرة لاحظتها بليد (1955) في الخلايا الحيوانية. عزل الريبوسومات واكتشف الحمض النووي الريبي منها ولهذا السبب يطلق عليه أيضًا RNP أو جزيئات بروتين الريبونات أو حبيبات بالاد. [كلود] عيّنهم بما أنّ [ميكروسوم] غير أنّ عيّنت اسم [ريبوسم] كان بروبرت في 1958.
حادثة:
فهي موزعة عالميا في جميع أنحاء المملكة الحيوانية والمملكة النباتية. بدائيات النواة أيضا ، تم العثور عليها. أنواع الخلايا الوحيدة الخالية من الريبوسومات هي RBC الثدييات. كثافة الريبوسوم لكل وحدة مساحة ثابتة إلى حد ما لأي نوع معطى. وهي عالية في الخلايا التي تخلق بروتين m نشط وانخفاض في الخلايا حيث تكون تخليق البروتين منخفض.
توزيع:
في الخلايا بدائية النواة ، غالبا ما تحدث الريبوسومات بحرية في السيتوبلازم. في الخلايا حقيقية النواة ، الريبوسومات إما تحدث بحرية في السيتوبلازم أو تظل متصلة بالسطح الخارجي لغشاء الشبكة الإندوبلازمية (ER).
عندما لا يتم ربطهم بالـ ER ، يطلق عليهم الريبوسومات الحرة. الريبوسومات الحرة بمثابة مواقع لتوليف البروتينات المطلوبة للحفاظ على دستور الانزيم للمصفوفة السيتوبلازمية.
طريقة العزل:
وعادة ما يتم عزل الريبوسومات عن الخلية بطريقة الطرد المركزي التفاضلي التي تستخدم فيها أجهزة الطرد المركزي التحليلية. يتم تحديد معامل الترسيب للريبوزومات من خلال التقنيات البصرية والإلكترونية المختلفة. يتم التعبير عن معامل الترسيب m وحدة Svedberg ، على سبيل المثال ، وحدة 'S'. يرتبط S بالحجم والوزن الجزيئي للجسيمات الريبوسومية.
عدد وتركيز الريبوسومات:
في جميع الخلايا التي تحتوي على شبكية endoplasmic ، يمكن ملاحظة عدد كبير من الريبوسومات. على سبيل المثال ، عند قاعدة خلايا الغدة ، في خلايا البلازما والكبد ، وفي جميع الخلايا النباتية والحيوانية سريعة النمو وفي البكتيريا ، يمكن أن ترتبط كمية من حمض الريبونوكلييك (RNA) بتركيز الريبوسوم.
في الخلايا الشبكية للأرانب تم العثور على ما يقرب من 100 ريبوسوم لكل، 3 ، والتي تقابل 1 × 10 5 جزيئات لكل خلية شبكية وحوالي. 5٪ من الحجم الإجمالي للكتلة الخلوية ، أو حوالي 20000 إلى 30000 خلية لكل خلية. ومع ذلك ، إذا تباطأ معدل تخليق البروتين بسبب الظروف الغذائية غير المواتية ، فإن عدد الريبوسومات يمكن أن ينخفض بشكل كبير في خلايا تخليق البروتين والبكتيريا.
هيكل ريبوسوم:
تتميز الريبوسومات بتناسقها في الحجم والتكوين من خلال مجموعة واسعة من الخلايا التي تمت دراستها فيها. ريبوسوم من النباتات والحيوانات أعلى هي المفلط ، الأجسام الشبه الكروية وقطرها ما يقرب من 250 درجة مئوية.
ومع ذلك ، فإن ريبوزومات البكتيريا أصغر إلى حد ما ، لأنها تحتوي على كميات أقل من البروتينات من ريبوسوم الحيوانات الأعلى. ومع ذلك ، في كمية من الحمض النووي الريبي في الجسيم ، يشبه الريبوسوم البكتيري جميع الريبوسومات الأخرى التي تمت دراستها في الوقت الحاضر.

في الدراسات المجهرية الإلكترونية يكشف التلوين السلبي عن شق ينقسم الريبوسومات إلى وحدة فرعية أكبر ووحدة فرعية أصغر. في E. coli يكون الجسيم الأكبر إلى حد ما على شكل "كوب" أو على شكل قبة (140 إلى 160 درجة مئوية) وصغيرًا يشكل "غطاء" (90 إلى 110 درجة مئوية) يتم تطبيقه على السطح المسطح للآخر (Huxley) وزوبي 1960). في الحيوانات والنباتات الأعلى ، ظهر أن الريبوسوم مرتبط بالشبكة الإندوبلازمية بواسطة الوحدات الفرعية الكبيرة.
البنية الدقيقة للريبوسوم معقدة للغاية ولم يتم توضيحها بالكامل بعد. بما أن الريبوسومات مسامية للغاية ورطبة ، فإن ال Rروتين RNA والبروتين ربما يكونا متشابكين داخل الوحدتين الفرعيتين ، في أقسام ، ملطختين بأيونات اليورانيل (RNA selective stain) ، كل ريبوسوم يظهر كجسم على شكل نجمة بأربعة إلى ستة أذرع مزروعة على محور كثيف. تظهر الوحدة الفرعية 50S المعزولة لعناوين Bacillus كجزيء مدمج من 160 إلى 180 درجة ، لها وجه خماسي ، في وسطها مساحة دائرية من 40 إلى 60 درجة (Nanninga، 1967).
تم وصف قلب شفاف شفاف الإلكترونات ، والذي يكون ملطخًا بالسالب في الريبوسومات ، مع منطقة إلكترون كامد ، في المقتطفات الكبيرة. الوحدة الفرعية 40S غير منتظمة وتميل إلى أن تقسم إلى جزأين متصلين ببعضهما بسمك 30 إلى 60 درجة.
الوحدة الفرعية الريبوسوم:
خاصية أخرى هي مشتركة بين جميع الريبوسومات في تركيبها الوحشي وثابتهم الترسيبي عن طريق تنبيذ فائق. على أساس ثابت الترسيب هناك نوعان رئيسيان من ريبوسوم.
تلك من البكتيريا لديها عادة معامل 70S (وحدة Svedberg) المقابلة للوزن الجزيئي 2.7 × 10 6 . والأخرى هي ريبوسوم الخلايا حقيقية النواة (الخلايا المنواة ، إما نباتية أو حيوانية) ، حيث يمتلك ريبوسوم ثابت ترسب 80S بوزن جزيئي يبلغ حوالي 4 × 10 6 daltons.

تتطلب المنطقتان البنيويتان المذكورتان أعلاه (الكوب وغطاء) من الريبوسومات تركيزًا منخفضًا من أيونات Mg ++ (0.001M) للتماسك الهيكلي. يمكن تنظيف الريبوسومات عن طريق إزالة أيونات Mg ++ لإعطاء جزيئين أصغر واحد 2/3 و 1/3 أخرى من كتلة الريبوسوم الأصلي الأصلي. في الواقع ليست سوى نفس الوحدات الفرعية التي لوحظت بمساعدة من المجهر الإلكتروني.
يشار إلى هذه الوحدات الفرعية من ثوابت الترسيب الخاصة بهم. تمتلك وحدة واحدة من الريبوسوم ثابت ترسب من 80S أو 70S كما ذكر من قبل ، في حين أن الوحيدات 2/3 لديها ثابت لترسيب 60S أو 50S و 1/3 subunit 40S أو 30S على التوالي. ترتبط كلتا الوحدتين معا من خلال أيونات المغنيسيوم التي تتفاعل مع مجموعات فوسفوديستر من الحمض النووي الريبي.
يحتوي الجسيم المشبع بالمغنيسيوم بالكامل على أيونات Mg ++ لكل ثلاث مجموعات فوسفودايتر. إزالة ثلث محسوب من هذه الأيونات المغنيسيوم يؤدي إلى انشقاق من 80S لتشكيل 60S و 40S (الوحدات الفرعية).
إزالة مزيد من Mg + + يؤدي في حالة من البازلاء والخمائر الريبوسوم على الأقل في مزيد من الانقسام تحرير ما هو على ما يبدو 1/6 حدة فرعية. غير أن هذا الانقسام الإضافي ، على النقيض من ذلك إلى الوحدات الفرعية 2/3 و 1/3 ، لا يمكن عكسه بمعنى أن استعادة أيونات المغنيسيوم ، لا يؤدي إلى استعادة جزيئات 80S.
إذا زاد تركيز Mg ++ بمقدار عشرة أضعاف ، فسيتم الجمع بين اثنين من ريبوسوم لتشكيل "ديمر" مع ضعف الوزن الجزيئي للريبوسوم الفردي. يمكن تحويل الثنائيات مرة أخرى إلى ريبوسوم عن طريق خفض تركيز Mg ++ .

كشفت المشاهدات المجهرية الإلكترونية في وقت مبكر لأقسام الخلايا أن الريبوسومات كانت ترتبط في كثير من الأحيان في مجموعات تشكل في بعض الأحيان نمط متكرر. لم يكن حتى عام 1962 أن اكتشف وظيفة هذه polyribosomes ، أو polysomes ، في تخليق البروتين (وارنر وريتش ، 1962).
بعد معالجة الخلايا الشبكية مع الأحماض الأمينية المسمى C 14 واستخدام طرق لطيفة للاضطراب ، وجد أنه بالإضافة إلى ثابت الترسب النموذجي للريبوسوم المفرد (80S) كانت توجد وحدات أكبر.
يختلف ثابت الترسب لهذه الجسيمات من 108S إلى 170S أو أكثر ؛ هذا يتوافق مع polyribosome من خمس وحدات (البنتامر). وقد تم تأكيد ذلك بواسطة الفحص المجهري الإلكتروني بأن حوالي 75٪ من الريبوسومات التي تظهر ذروتها في 170S ، تم تقديمها على هيئة pentameres.
خيوط رقيقة ، تفسر في mRNA ، حوالي 150 A °. قد يختلف عدد الريبوسومات في polyribosomes اختلافًا كبيرًا ويبدو أنه مرتبط بطول الرنا المرسال الذي ينبغي "قراءة" في عملية الترجمة.
في E. coli وفي الخلايا من جنين الفرخ ، تمت ملاحظة polyribosomes المكونة من حوالي 50 وحدة {Rich 1967}. قد تكون polyribosomes خالية في السيتوبلازم أو مرتبطة بأغشية الشبكة الإندوبلازمية.
بالنسبة للـ polyribosomes الحرة ، تم افتراض تكوين حلزوني مع الوحدات الفرعية الصغيرة التي تم ترتيبها حول المحور المركزي والوحدات الفرعية الكبيرة التي يتم التخلص منها في المحيط الخارجي. في أقسام من الخلايا المختلفة تم رصد polyribosomes مجموعة دين حلزونية (Weiss و Grover ، 1968). من المفترض أنه في polysome يقع mRNA بين اثنين من وحدات فرعية من الريبوسوم.
أنواع الريبوسومات :
الريبوسومات هي من نوعين أساسيين ، 70S و 80S الريبوسوم. وهكذا يشير S إلى وحدات Svedberg. في الواقع هو معامل الترسيب الذي يظهر مدى سرعة رسوبيات الخلايا العضوية في نابذة فائقة السرعة. معاملات الترسيب ليست مضافة.
تم العثور على الريبوسومات 80S في حقيقيات النوى (الكائنات الحية التي تحتوي خلاياها على نويات حقيقية محاطة بالمظاريف النووية) مثل الطحالب والفطريات والنباتات والحيوانات. يتكون الريبوسوم 80S من الحيوانات من الوحيدات 60S كبيرة ووحدة فرعية صغيرة 40S.
70S الريبوسومات هي أصغر نسبيا وتوجد في بدائيات النواة (الكائنات الحية التي لا يحدها الحمض النووي عن طريق غلاف نووي) على سبيل المثال البكتيريا. يتكون الريبوسوم 70S من وحدة فرعية كبيرة 50S ووحدة فرعية صغيرة 30S.
الريبوسومات الموجودة في الميتوكوندريا والبلاستيدات الخضراء من حقيقيات النواة هي أقرب إلى الريبوسومات بدائية النوى بدلا من ريبوسوم حقيقيات النواة 80S. وتحتوي الميتوكوندريا الفقارية ، على سبيل المثال ، على ريبوزومات 55S ، لكل منها وحدة فرعية كبيرة 40S ووحشية صغيرة 30S. يتم تقريب معاملات الترسيب 80S و 70 S و 55S من القيم. قد تكون قيم S الفعلية في الكائنات الحية أعلى أو أقل قليلاً.


التركيب الكيميائي :
المكونات الرئيسية للريبوسوم هي RNA والبروتينات. الدهون هي غائبة تماما أو موجودة في آثار. وتمتلك ريبوسومات الإشريكية القولونية ما يقرب من 60-65٪ من الرنا و 35-40٪ من البروتين من وزنها. يبين الجدول 6.3 كمية تقريبية من الحمض النووي الريبي والبروتين في أنواع مختلفة من الريبوسومات.
RNA Ribosomal يختلف في حجم ومحتويات قاعدة من الحمض الريبي النووي النقال ومن فئات الحمض النووي الريبي الأخرى لمعظم الخلايا. تم العثور على نوعين من الرنا في جميع ريبوسوم. فهي جزء لا يتجزأ ولا يمكن إزالتها بسهولة. الحمض النووي الريبي من جزيئات الفئران يحتوي أساسا على القواعد المشتركة الأدينين ، الجوانين ، السيتوزين واليوراسيل مع كمية صغيرة من pseudouridine ، يتم العثور على القواعد المعتادة التي تحدث في الحمض النووي الريبي القابلة للذوبان في الجسيمات RNA فقط في كمية صغيرة جدا.
الجدول 6.2 الاختلافات بين 70s و Ribosomes:
حضور | في البكتيريا بدائيات النوى ) | في حقيقيات النواة (الطحالب والفطريات والنباتات والحيوانات العليا) |
معامل الترسيب | 64S-72S (معدل 69S) | 79-85S في الفطريات ، 80S في الثدييات. |
بحجم | أصغر نسبيًا | أكبر نسبيا |
الوزن الجزيئي. | 3 × 10 6 | 4-5 × 10 6 |
الوحدات الفرعية | صغير 30S و large50S | صغير 40S و large60S. |
RNA | 3 جزيئات من الحمض النووي الريبي RNA16S في 30S الوحيدات ، 23S و 5 S RNA في الوحيدات 50S. | 4 جزيئات من الحمض النووي الريبي RNA16S-18S في 40S الوحيدات ؛ 25-29S ، 5.8S و 5 S RNA في الوحيدات 60S. |
م. من الحمض النووي الريبي | 16S RNA-550،00023 S RNA-1،100،0005S RNA-40،000 | 18S RNA-700،00028S RNA-1، 700، 0005.8S RNA 51،000 5S RNA -29،000. |
عدد البروتينات | 21 (S1-S21) في الوحيدات الصغيرة 34 (S1-L34) في الوحيدات الكبيرة مجموع في بدائيات النواة: 50-60 بروتين. | 33 في subunit49 الصغيرة في الوحيدات الكبيرة مجموع "في البروتينات eukaryotest70-80 |
متوسط م. من البروتين | 18000 | 21000 |
لا من الأحماض الأمينية | 8000 | 16000 |
نسبة RNA-Protein | 2.1 | 1: 1 |

أحماض Anino:
تم تحديد تكوين الأحماض الأمينية من بروتين حامض ثلاثي كلور حمض غير قابل للذوبان من RNP الكبد من قبل كرومبتون وبترمان (1959). كانت الأحماض الأمينية العطرية والكبريتية متواجدة بكميات قليلة جداً ، في حين أن الليوسين والأرجينين كانا أعلى من 10٪. يتشابه تكوين كل من شبكيات الأرانب الشبكية و RNPs من البسلة بشكل ملحوظ.
وقد أنتج المستخلص الهيدروكلوريكي من RNP المنقى جزئياً RNP أرجينين (Butter، 1960) ، ولكن ما إذا كانت هذه الأحماض الأمينية مشتقة من البروتينات نفسها لم يتم تحديدها بعد.
بروتين:
يحتوي البروتين على سلسلة خطية من الأحماض الأمينية. مماثلة في تكوين الريبوسومات من الأحماض الأمينية للبروتين الريبوسومي هو من أصل مختلف بشكل ملحوظ ، وربما يكون مختلفا تماما عن البروتين المكون من الريبوسوم.
لذلك ، من الشائع التمييز بين البروتين الهيكلي الريبوسومي ، فإن سلسلة الببتيد المتنامية من الإنزيم الذي يتم تصنيعه بواسطة الريبوسوم الذي يرتبط البروتين بالحمض الريبي النووي الريبوزي بواسطة الرابطة الهيدروجينية واضح من حقيقة أن انفصال الاثنين يتم بسرعة وبسرعة عن طريق الكواشف يهاجم الروابط الهيدروجينية على سبيل المثال بروميد الجوانيديوم.
تم إجراء التجارب بواسطة يين وبوك (1960) ، اللذان حصلا على بروتين ريبوزومي مستقر. حصل واتسون (1960) على بروتين E.coli ، ويشير تحليل مثل هذه المجموعة إلى أن الوزن الجزيئي لكل وحدة فرعية يبلغ حوالي 30،000.
البروتينات الأنزيمية الريبوسومية:
معظم البروتينات الريبوسومية تعمل كأنزيمات وبالتالي تحفز تخليق البروتين. تبدأ بروتينات البدء IF 1 و IF2 و 1F3 بعملية تخليق البروتين في حين أن بروتينات النقل (G-factor، Ts-factor) تساعد في نقل الريبوسومات فوق mRNA ونقل رواسب t-RNA من موقع الريبوسوم إلى موقع آخر. .
يساعد الانقلاب الببتيدى 1 الآخر في تحويل سلاسل الببتيد إلى aminoacy1-tRNA والإنزيمات الأخرى لإنهاء سلسلة polypeptide المكتملة.
نتيجة لغسيل الريبوسوم مع NH 4 CI وإخضاعها لعمود كروماتوغرافي عمود ، عزل Ochoa وزملاء العمل (1960) فوق العوامل في تخليق البروتين. فيما بينها ، ترتبط ثلاثة عوامل البدء - IF1 و IF2 و IF3 - بشكل فضفاض بالوحدات الفرعية 30S. إن عامل IF1 عبارة عن بروتين أساسي له وزن جزيئي 9200 daltons.
وتشارك في ربط F-met-tRNA. عامل IF2 هو أيضا بروتين من مول. بالوزن. 8000 daltons ومجموعة تحتوي على SH تساعد في الارتباط بـ GTP. عامل البروتين الثالث ، IF3 ، لا يتطلب GTP ويشارك في ربط مرنا إلى وحدات جزئية 30S.
وهو بروتين أساسي له وزن جزيئي يبلغ 30 ألف daltons. قد يعمل IF3 أيضًا كعامل تفكك لـ 70S ريبوسوم. Ochoa وآخرون. (1972) قد أبلغت عن تدخل عامل (ط) في البكتيريا E.coli. هذه العوامل ترتبط بعامل IF3 ، وتغير خصوصيتها وبالتالي تنظم ترجمة الرسالة الجينية في البداية.
عوامل الاستطالة ضرورية لإطالة سلسلة polypeptide. هذه هي EFG (وتسمى أيضا عامل G أو translocase) وعامل EFT. كما هو موضح في وقت سابق EFG أو عامل G تشارك في عملية تحويل mRNA. في E. coli ، تتكون من سلسلة بولي ببتيد مفردة لها جزيء. بالوزن. من 72 ، 00 daltons. EFG + GTP يشجع على الانتقال من peptidyl-tRNA الممدود حديثا.
هناك عامل آخر لتحويل الأموال الإلكترونية له نوعان من البروتينات وهما Tu (درجة الحرارة غير مستقرة) و Ts (درجة حرارة ثابتة). يقوم عامل EFTU + GTP بتشكيل aminoacyl-tRNA المعقدة قبل ربطه بالموقع المتقبل للريبوسوم الذي يتم تحفيزه بواسطة معامل Ts. علاوة على ذلك ، تحتوي الوحدة الفرعية الريبسيمية 50S على مركب إنزيم ببتيد إنزيم أو ترانسفيراز peptdyl المرتبط بتكوين رابطة الببتيد. تقوم عوامل الإنهاء R1 و R2 بإطلاق بروتينات تساعد في تحرير سلسلة polypeptide.
بيوجينيسيس من ريبوسوم:
يحدث التكوُّن الحيوي للريبوزومات في الخلية بدائية النواة البكتيرية داخل السيتوبلازم بسبب غياب النواة. تنشأ rRNAs من الكودونات المحددة للجينوم أو الحمض النووي الريبوزومي (rDNA).
في الخلايا حقيقية النواة ، تكون عملية التكوُّن الحيوي للريبوزومات معقدة وتحدث داخل النواة. واحد من الكروموسومات في مجموعة لديها منطقة تنظيمية محددة من النواة ، والتي تحتوي على جزيء الحمض النووي الريبي النووي الذي هو مقدمة لكل من الحمض الريبي النووي الريبي 28S و 18S. عملية التحويل من 45S-RNA إلى 28S و 18S rRNA موضحة أدناه:
جزيئات RNA النواة 45S هي ميثليته (-CH ، المجموعة مضافة). تصبح هذه الجزيئات الميثيلية من الحمض النووي الريبي النووي الجزيئي 45S مرتبطة بالبروتينات الضرورية الموجودة في النواة التي تشكل جزيئات جزيئات ريبونسيتورين (RNP) 80S. هذه RNP 80S مع الحمض النووي الريبي 45S الجزيئي تقسيمها إلى 32S و 18S rRNA من خلال عدة خطوات وسيطة ، والتي يتم فقدان جزء غير الميثيل من الجزيئات. جزيء 18S مع جزيئات البروتين التي تنتقل على الفور إلى السيتوبلازم. يبقى الحمض الريبي النووي الريبي (32S) في داخلها النواة لبعض الوقت ويحصل على شق في rRNA 28S.
يتم تصنيع الرنا الريباسي 5S rRNA خارج النواة ، وتقع الجينات بجوار المنطقة المنظِّمة للنواة في الكروموسوم.

إن الـ 18S rRNA مع بروتيناته تترك النواة من خلال nucleopore وتخرج إلى السيتوبلازم ، حيث تترابط مع البروتينات إلى وحدات صغيرة (40S) من الريبوسوم. ويترك الـ 28S rRNA أيضًا النواة وتدمج مع rRNA 5S والبروتينات المكونة لوحدة 60S.
يحدث تخليق البروتين الريبوسومي جزئيا داخل النواة وجزئيا داخل السيتوبلازم. يتم تجميع البروتينات في تجمع السيتوبلازم في النواة ليتم استخدامها في جسيمات جزيئات ريبونوكليروتين (RNP).
ريبوسومال رنا :
يحتوي الريبوسوم 70S على ثلاثة أنواع من rRNA ، أي ، rRNA 23S ، و rRNA 16S و 5S rRNA. يحدث الرنا الريباسي 23S و 5S في وحدة فرعية الريبوسوم 50S الأكبر ، في حين يحدث الـ rRNA 16S في الوحدة الفرعية الريبوسومية الأصغر 30S. ويتكون الرنا الريباسي 23S من 3200 من النيوكليوتيدات ، ويحتوي 16S rRNA على 1600 نيوكليوتيدات و 5S rRNA تتضمن 120 نيوكليوتيد فيها (Brownlee، 1968، Fellner، 1972).
يحتوي الريبوسوم 80S أيضًا على ثلاثة أنواع من rRNA ، أي rRNA ، أي 28S rRNA ، و rRNA 18S و rRNA 5S. تحدث الحمض الريبي النووي الريبوزي 28S و 5S في الوحيدات الريبوسومية الكبيرة 60S ، في حين يحدث الـ rRNA 18S في الوحدة الفرعية الريبوسومية الأصغر 40S.
يحتوي الـ 28S rRNA على الوتر الجزيئي بنسبة 1.6 x 10 6 daltons وجزيئه مزدوج الضفيرة وله قواعد النيتروجين في أزواج. الحمض الريبي النووي الريبي 18S لديه الوزن الجزيئي 0.6 * 10 6 daltons. يحتوي جزيء rRNA 5S على شكل ورقة البرسيم وطول يساوي 120 نوكليوتيد (Forget and Weissmann، 1968).
المكونات الأخرى :
محتويات المعدن من ريبوسوم هي أيضا مسألة صعبة. ليس هناك شك في أن المغنيسيوم هو رئيس الأيونات من الناحية الفسيولوجية ، لأنه موجود في تركيزات عالية في ريبوسوز الكبد الفئران. وهو الأكثر نشاطًا في الحفاظ على بنية 80S (هاميلتون و Petermann ، 1959) حيث أنه ضروري لتضمين الأحماض الأمينية في المختبر (Rendi and Hultin، 1960). المعادن الأخرى الموجودة في الريبوسومات هي الكروم والمنغنيز والنيكل والحديد والكالسيوم (Walker and Valle، 1958؛ Tso 1958).
وظيفة ريبوسوم :
من الحقائق المعروفة جيداً أن الريبوسومات هي مصانع لتصنيع البروتين في الخلية ، لكن في الواقع لا يمكن أن يشارك ريبوسوم واحد في تخليق البروتين. كان معروفا منذ عام 1962 عندما تم نشر التقرير الذي أظهر أن الوحدات النشطة ليست ريبوسوم الفردية ، ولكن مجموعة من هذه الوحدات ، والتي يطلق عليها اسم polyribosome. تم نشر دور مفصّل لـ polyribosome في مجلة Scientific American ديسمبر عام 1963 بواسطة Gric and Hall. هناك الكثير من الأدلة على أن الريبوزوم متشابه وأنهما قابلتان للتبادل على الأقل.

في حالة الهيموغلوبين مع سلسلة من الأحماض الأمينية 150 ، تم وصف العدد الأكثر شيوعا من ريبوسوم وما يصل إلى 40 ريبوسوم. لذلك يمكن أيضا استخدام polysome اسم في مكان polyribosome. يتكون البروتين من سلسلة خطية من الأحماض الأمينية. قد تكون السلسلة قصيرة أو طويلة في كلتا الحالتين تسمى سلسلة ببتيد. يمكن طي البولي ببتيد بطريقة معينة في كثير من الأحيان لتكوين بروتين معقد.
يتم فصل الريبوسومات في polysome بفجوة من 50-150 A 0 . وتكشف النتوءات الإيجابية من قبل Uracil-acetate أنه يتم توصيل الريبوسومات بواسطة خيط رفيع من 10-15A 0 في القطر ، وهو حول سمك خيط وحيد من RNA. من حجم الفجوة بين الريبوسوم ، فإن القياس الإجمالي للفجوة التي تحتوي على خمسة ريبوسوم ، حوالي 1500A 0 ، و reibosomes خمسة لديها فجوة بين الرايبوسومات من 50-150A ° لكل منهما.